
Экосистемы, 30: 84–94 (2022) https://ekosystems.cfuv.ru
![]()
УДК 581.524.2(470-924.71)
Морфологическая изменчивость Ambrosia artemisiifolia в условиях Предгорного Крыма
Омельяненко Т. З.1, Багрикова Н. А.2
- Южный филиал ФГБУ «Всероссийский центр карантина растений» Симферополь, Россия
o.tanya-work@yandex.ru
- Никитский ботанический сад – Национальный научный центр РАН Ялта, Республика Крым, Россия
Одним из наиболее широко распространённых инвазивных растений в России, демонстрирующих высокую экологическую пластичность, является североамериканский вид – Ambrosia artemisiifolia L. В работе представлены сведения о его распространении в Республике Крым. Проведен анализ изменчивости морфометрических параметров 12 ценопопуляций, изученных в 2021 году в различных экотопах Предгорного Крыма. Наибольшие значения морфометрических показателей отмечены в синантропном и рудеральном местообитаниях, где для сообществ характерен выраженный уровень антропогенной нагрузки. Наименьшие средние значения отмечены в синантропизированных сообществах. Наибольшей степенью изменчивости отличались количественные параметры. Минимальной изменчивостью характеризовались качественные параметры генеративной сферы. На основании корреляционного анализа установлено, что наибольшее число очень сильных и сильных связей (r>0,71) характерно для качественных параметров. На основании дискриминантного и кластерного анализов установлено, что наибольшее фенотипическое сходство характерно для ценопопуляций в синантропизированных и рудеральных местообитаниях. Кластерный анализ позволил объединить 12 ценопопуляций в 4 группы. В отдельный кластер выделяется ценопопуляция, описанная в синантропизированном разнотравном сообществе. Она отличается от других ценопопуляций наименьшими средними значениями морфометрических параметров.
Ключевые слова: инвазивное растение, карантинный вид, ценопопуляции, изменчивость, Предгорный Крым.
ВВЕДЕНИЕ
Попадание чужеземных видов растений в новые условия неизбежно влечет за собой ряд трудно прогнозируемых последствий. Среди них особую роль играют процессы борьбы с аборигенными видами за ресурсы, а также трансформация захваченного пространства, что создает менее благоприятные условия существования слагающих сообщества видов. Некоторые исследователи отмечали, что усиление антропогенного воздействия способствует плавному стиранию границ экологически разнообразных флор (Бурда, 1991; Тохтарь, Петрик, 1993; Тохтарь, 1996), а последующее внедрение агрессивных чужеземных видов ведёт к блокировке сукцессионных процессов и может стать причиной исчезновения доминирующих растительных сообществ (Шварц, 2004). Итогом подобного вмешательства является нарушение естественного хода флорогенеза.
Несмотря на богатый арсенал высокотехнологичных методов, применяемых при изучении чужеродных видов, стоит признать, что достоверно установить комплекс универсальных признаков, способствующих их успешной экспансии, крайне затруднительно. Иногда закономерности, которые лежат в основе успешной инвазии вида в одних условиях, могут оказаться совершенно неприменимы в иных. В связи с этим, особую актуальность представляет изучение изменчивости морфометрической структуры вида в разнообразных биотопах, что позволяет оценить его адаптационные возможности. Подобные исследования приближают к более точному прогнозированию инвазии и ее успешности, способствуют разработке комплекса эффективных мер, а также выбору информативных маркерных
ISSN 2414-4738 Published by V. I. Vernadsky Crimean Federal University, Simferopol
Морфологическая изменчивость Ambrosia artemisiifolia в условиях Предгорного Крыма
![]()
морфологических параметров, детерминирующих популяционную структуру вида в определенных условиях.
-
- России одним из наиболее широко распространённых инвазивных растений, демонстрирующих высокую экологическую пластичность, является североамериканский вид Ambrosia artemisiifolia L. В зависимости от условий обитания, растения способны проявлять широкий спектр биоморфологической изменчивости, семенной продуктивности и, как следствие, степени инвазивности. Амброзия полыннолистная входит в перечни карантинных видов ряда стран Азии (Иордания, Казахстан, Узбекистан), Европы (Азербайджан, Беларусь, Российская Федерация, Испания, Швейцария и Украина) и Африки (Египет). Вид включен в перечни ЕАЭС и ЕОКЗР (EPPO Global Database).
- «Биологической флоре Крыма» (Голубев, 1996) A. artemisiifolia отнесена к адвентивным видам и указана для предгорного, южнобережного Крыма, Керченского полуострова. Анализ литературных данных, гербарных образцов (YALT и Национального банка-депозитария живых систем «Ноев ковчег») и собственных полевых исследований показал, что в Крыму A. artemisiifolia распространена во всех природных зонах и отмечена в Симферопольском, Белогорском, Красногвардейском, Кировском, Нижнегорском, Ленинском, Первомайском, Черноморском, Сакском, Джанкойском, Бахчисарайском, Советском, Красноперекопском, Раздольненском административных районах, а также в пределах Ялтинского, Алуштинского, Судакского, Феодосийского, Евпаторийского городских округов Республики Крым, а также на территории города федерального значения Севастополь (Беляева, Пирогова, 2020; Плантариум, 2022; Global Biodiversity Information Facility, 2022; iNaturalist, 2022).
На территории Крымского полуострова амброзия отмечается в составе настоящих степей
- предгорных лесостепей и имеет третий статус инвазионности. Таким образом, она относится к группе чужеродных видов, которые заселяют нарушенные местообитания, и, натурализуясь в них, способна внедряться как в естественные, так и полуестественные растительные сообщества (рис. 1) (Багрикова, Скурлатова, 2021).
Цель работы – изучить морфологическую изменчивость A. artemisiifolia в различных эколого-ценотических условиях Предгорного Крыма.
МАТЕРИАЛ И МЕТОДЫ
Изучение 12 качественных и количественных параметров морфоструктуры вида проводилось на 25 средневозрастных растениях по общепринятой методике (Голубев, 1962) в 12 ценопопуляциях в различных эколого-ценотических условиях. Для анализа применялся метод, предложенный С.А. Мамаевым (1985), по которому на основании показателя коэффициента вариации (CV, %), выделяются следующие уровни изменчивости признаков:
- 7 % – очень низкий; 8–12 % – низкий; 13–20 % – средний; 21–40 % – высокий; более 40 % – очень высокий. Использован индекс фитоценотической пластичности (Ip), который рассчитывается по формуле:
![]()
где: Ip – индекс фитоценотической пластичности; А и В – максимальные и минимальные средние значения признака соответственно (Злобин, 1989).
Взаимосвязь между параметрами оценивалась по коэффициенту корреляции, значения которого позволили подразделить степень связи на 4 основные группы: r>0,8 – очень сильная связь; r=0,71–0,8 – сильная связь; r=0,61–0,7 – умеренная связь; r=0,5–0,6 – слабая связь (Пархоменко, Кашин, 2011). С целью установления фенотипического сходства и/или различия ценопопуляций применялся дискриминантный анализ с учетом расстояния Махаланобиса. Кластерный анализ проводился с использованием иерархической классификации и
85
Омельяненко Т. З., Багрикова Н. А.
![]()
кластеризации методом k-средних с применением метода Варда и учетом Евклидова расстояния. Статистическая обработка выполнена с помощью пакетов программ MS Excel 10
- STATISTICA 10.
Исследование проводилось в Симферопольском и Белогорском административных районах, входящих в юго-западный и центральный предгорные агроклиматические районы Крыма (Опанасенко и др., 2015).
ЦП 1. 45.067234 N, 34.452202 E. Белогорский район, сельское поселение Ароматное. Синантропное сообщество на окраине поля зерновых, где сформировались монодоминантные заросли амброзии. Общее проективное покрытие (ОПП) – 100 %, A. artemisiifolia – 95 %.
ЦП 2. 45.062491 N, 34.496496 E. Белогорский район, сельское поселение Зеленогорье. Сегетальное сообщество на убранном поле зерновых. У многих растений при уборке урожая был обрезан главный побег, в связи с чем сформировались боковые побеги. По осям мужских соцветий развивались дополнительные ответвления с расположенными на них корзинками и завязями. ОПП – 80 %, A. artemisiifolia – 20–30 %.
ЦП 3. 45.07693 N, 34.459225 E. Белогорский район, сельское поселение Зеленогорье. Синантропное сообщество вблизи заброшенного поля зерновых. ОПП – 100 %, A. artemisiifolia – 50 %.
ЦП 4. 45.032225 N, 34.345765 E. Белогорский район, сельское поселение Крымская Роза. Ценопопуляция расположена в пределах синантропизированного сообщества в составе сосняка, внутри которого был отмечен крупный перекопанный участок, где были удалены все древесные растения (рис. 1f). В результате этого по периферии антропогенно нарушенного участка, произрастает лесное сообщество, в состав которого внедрились синантропные представители. Единичные особи амброзии распространились вглубь соснового леса на расстоянии до 150 м от основной части ценопопуляции. ОПП – 90 %, A. artemisiifolia – 45– 50 %.
ЦП 5. 45.051978 N, 34.593475 E. г. Белогорск, ул. Гагарина. Рудеральное сообщество у гаражного кооператива (рис 1e). ОПП – 90 %, A. artemisiifolia – 75 %.
ЦП 6. 44.945249 N, 34.226144 E. Симферопольский район, рудеральное местообитание вдоль трассы Трудовое – Денисовка, вблизи Таврической ТЭС. Ценопопуляция представлена очень мелкими особями. ОПП – 90 %, A. artemisiifolia – 15–20 %.
ЦП 7. 45.13868 N, 34.600539 E. Белогорский район, с. Вишенное. Рудеральное сообщество у частного домовладения, ОПП – 90 %, A. artemisiifolia – 50 %.
ЦП 8. 45.067551 N, 34.569867 E. г. Белогорск. Рудеральное сообщество на обочине трассы «Таврида», вблизи ул. Муарема Мартынова. Ценопопуляция представлена относительно мелкими особями. У большинства растений отсутствовал главный побег, выражены побеги 2– го порядка. Для некоторых особей отмечены многочисленные женские цветки, мужские корзинки практически отсутствовали. ОПП – 55 %, A. artemisiifolia – 10–15 %.
ЦП 9. 45.06482 N, 34.476316 E. Белогорский район, с. Русаковка, вблизи лесного массива Кара-Куш (рис 1a). Синантропное сообщество между краем поля подсолнечника и трассой «Таврида». ОПП – 100 %, A. artemisiifolia – до 70–90 %.
ЦП 10. 44.59817 N, 34.74348 E. Симферопольский район, трасса 35А-002 (поворот с Симферопольской объездной дороги (Мирное – Дубки) в сторону пос. Айкавана). Рудеральное местообитание. У многих растений были скошены побеги, при этом у части растений отсутствовали мужские корзинки, а на их месте выявлены многочисленные женские цветки. ОПП – 85 %, A. artemisiifolia – 40 %.
ЦП 11. 45.06482 N, 34.476316 E, Белогорский район, А-291, 169-й километр. Синантропное сообщество между краем поля подсолнечника и трассой «Таврида». У многих особей были скошены главные побеги. ОПП – 90–100 %, A. artemisiifolia – до 80 %.
ЦП 12. 45.027827 N, 34.426989 E. Белогорский район, сельское поселение Зеленогорье, дорога в с. Межгорье. Синантропизированное разнотравное сообщество, используемое для сенозаготовки. ОПП – 80 %, A. artemisiifolia – до 20 %.
86
Морфологическая изменчивость Ambrosia artemisiifolia в условиях Предгорного Крыма

Рис. 1. Ambrosia artemisiifolia в различных экотопах Предгорного Крыма
| a | – | синантропное сообщество между краем поля подсолнечника и трассой | «Таврида»; |
| b | – | синантропное монодоминантное сообщество между с/х угодьями Белогорского района; | |
| c | – | единичные особи Ambrosia artemisiifolia в границах городской территории г. | Белогорска; |
d – сегетальное сообщество в Симферопольском районе; e – рудеральное сообщество около гаражного кооператива Белогорского района; f – синантропизированное сообщество на окраине соснового леса Белогорского района.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Наибольшие значения качественных (высота растения, длина и ширина листа, длина мужского верхушечного соцветия и диаметр мужской корзинки) и количественных (количество листьев и плодов на одно растение) морфометрических параметров отмечены для ЦП 7, описанной в рудеральном сообществе у частного домовладения, где ОПП – 90 %, а на A. artemisiifolia приходится 50 %. В ЦП 9 в синантропном сообществе между краем поля подсолнечника и трассой «Таврида» наибольшие значения характерны для диаметра побега, количества ветвлений и количества мужских корзинок в соцветии. Для ЦП 3 в синантропном сообществе с ОПП – 100 %, из которых на A. artemisiifolia приходится 50 %, установлено, что такие параметры, как количество мужских корзинок в соцветии, длина и ширина плода, отличаются высокими показателями (табл. 1).
Значения некоторых параметров заметно превышают средние показатели, представленные в таблице 1. Например, в ЦП 9 отмечались наибольшая высота – 138 см и
87
Омельяненко Т. З., Багрикова Н. А.
![]()
наибольшее количество листьев – более 1500 шт., а максимальное количество ветвлений было выявлено в ЦП 7 – 41 шт. Максимальное значение семенной продуктивности было характерно для некоторых особей в ЦП 9 и достигало 12760 шт.
Таким образом, наибольшие значения морфометрических показателей отмечены в синантропных, в том числе рудеральных сообществах Белогорского района с выраженным уровнем антропогенного пресса.
Практически все параметры особей ЦП 12, произрастающих в составе синантропизированного сообщества, где при ОПП 80 % на A. artemisiifolia приходится до 20 %, характеризуются наименьшими средними значениями. Можно предположить, что при высокой плотности травостоя представители семейств Poaceae и Fabaceae в этом сообществе за счет своей засухоустойчивости и выраженной конкурентной способности за разнообразные ресурсы среды, препятствуют внедрению и развитию A. artemisiifolia. Помимо этого, причинами миниатюризации особей амброзии могут являться сухость почв и бедность субстрата.
При сравнении морфометрических параметров растений амброзии в разных регионах Российской Федерации выявлены некоторые особенности.
-
- условиях Предгорного Крыма в 2009–2010 годах высота растений изменялась от 24,2±0,9 до 99,5±0,3 см (Жалдак, 2011), в рамках настоящего исследования в 2021 году средние значения варьировали в пределах от 18,8±1 до 96,9±7,1 см, что свидетельствует о стабильности указанного показателя.
- Приморском крае высота растений достигает 2 м, при этом средние значения варьируют от 1,0 до 1,2 м (Кудрявцева и др., 2018), что значительно превышает значения, полученные в условиях Предгорного Крыма. В условиях Республики Башкортостан в с. Ермолаево высота растений в среднем составляла 71,2±9,6 см (Абрамова, 2014), что соответствует значениям, описанным в синантропизированном и рудеральном сообществах Предгорного Крыма. В Кабардино-Балкарской Республике высота растений варьировала в зависимости от условий произрастания: в нарушенных рудеральных сообществах достигала всего 10,2±4,1 см; в естественных луговых фитоценозах изменялась от 12,2±3,1 до 48,4±12,3 см; в полуестественных фитоценозах – от 20,6±1,9 до 82,4±6,4 см; на сорных местах
– от 56,2±5,2 до 138,6±1 см (Чадаева и др., 2018). На основании того, что в синантропизированном сообществе Предгорного Крыма наименьшая высота растений составляла 10,2 см, а в синантропном сообществе высота самых крупных особей достигала 138 см, можно сделать вывод о том, что диапазон указанного параметра в Предгорном Крыму сходен с размахом, указанным в сорных и рудеральных сообществах Кабардино-Балкарской Республики.
Средние значения высоты растений в рудеральных и синантропных условиях Предгорного Крыма, в целом, соответствуют средним показателям высоты особей, описанных в Московской области, которые варьируют от 78 до 105 см (Петрова, 2019).
-
- Симферопольском и Белогорском районах в 2021 году размах длины и ширины листа находился в сравнительно широких пределах: средние значения длины листа варьировали от 3,1 до 13,3 см, ширины – от 1,7 до 8,1 см. В приведенный диапазон попадают и результаты, полученные в условиях Предгорного Крыма в 2009–2010 годах: длина листа – от 4,7±0,6 до 6,9±0,9 см, ширина – от 3,5±0,2 до 5±0,3 см (Жалдак, 2011). В Московской области указанные параметры также менялись в более узких пределах: длина листа – от 9 до 11 см, ширина – от 8 до 10 см (Петрова, 2019).
Морфометрические значения плодов амброзии в Предгорном Крыму (длина – от 3,3±0,1 до 4,5±0,2 мм; ширина – от 1,9±0,1 до 2,3±0,2 мм) несколько превышают показатели, установленные для плодов, собранных в Белгородской области России, где длина варьировала от 3,42±0,12 до 3,64±0,09 мм; ширина – от 1,9±0,06 до 2,16±0,06 мм (Тохтарь, 2012). При этом,
полученные в 2021 году данные уступают значениям, описанных в более ранних исследованиях в Предгорном Крыму, где длина плода достигала 4,8±0,05 мм (Жалдак, 2011). Однако в 2009–2010 годах семенная продуктивность вида достигала 371,4±10,1 шт., тогда как в 2021 году средние значения данного показателя варьировали от 38,8 до 4036,2 шт. Таким
88
Морфологическая изменчивость Ambrosia artemisiifolia в условиях Предгорного Крыма
![]()
образом, при сравнительно сходных эколого-ценотических условиях и морфометрических показателях вида, количественные репродуктивные параметры в 2021 году достигали значительно более высоких значений.
На основании проведенного анализа можно сделать заключение, что A. artemisiifolia демонстрирует широкий диапазон значений морфометрических параметров, как в сравнительно сходных условиях южных регионов РФ, так и в более северных районах.
-
- рамках настоящего исследования, во всех изученных ценопопуляциях максимальный размах варьирования и очень высокая изменчивость характерны для количественных параметров: количества ветвлений (31–97 %), количества листьев (36–97 %) и количества плодов на одно растение (57–99 %), которые относятся к наиболее адаптивным и пластичным. Наименьший размах варьирования отмечен для ширины листа (19–30 %), что свидетельствует
- сравнительной устойчивости этого параметра в разнообразных экотопах.
Высокие значения коэффициента вариации (CV – от 17 до 54 %) выявлены для высоты растения, диаметра побега, длины и ширины листа, длины мужского соцветия, а также количества мужских корзинок. Средние значения (2–36 %) характерны для диаметра мужской корзинки, низкие (0–19 %) – для ширины плода, очень низкие (0–16 %) – для длины плода (табл. 1).
Полученные данные подтверждают результаты анализа фитоценотической пластичности (Ip=17–99 %, в среднем – 69 %). Высокие индексы пластичности отмечались для всех параметров, кроме диаметра корзинки, длины и ширины плода. На основании этого можно сделать вывод, что качественные признаки генеративной сферы являются достаточно автономными и наименее чувствительными к изменениям среды, конкурентной нагрузке и антропогенному прессу. Они обусловлены преимущественно диапазоном генетических возможностей и не отражают общего состояния вида как восприимчивой системы.
Анализ изменчивости показал, что максимальное число наибольших значений коэффициента вариации выявлено в ЦП 7 и ЦП 8, описанных в рудеральных сообществах. Для ЦП 7 наибольшие показатели изменчивости выявлены для таких показателей как диаметр побега, количество листьев, а также длина и ширина листа; в ЦП 8 – для количества ветвлений и плодов, что обусловлено присутствием особей с обрезанными главными побегами.
Для ЦП 12, имеющей наименьшие значения морфометрических показателей, отмечается выраженная изменчивость для количества ветвлений, длины мужского соцветия, количества мужских корзинок и плодов. Это свидетельствует о том, что в экстремальных условиях вид проявляет выраженную гетерогенность элементов размножения, что увеличивает вероятность формирования жизнеспособных генеративных структур. В ЦП 3, описанной в синантропном сообществе, наибольшее значение коэффициента вариации (CV) установлено для высоты растения, которая варьировала от 27 до 88 см. Можно предположить, что монодоминантность сообщества способствовала беспрепятственному формированию нескольких генераций. Однако особи, которые развивались в более поздние сроки, не успели нарастить достаточную биомассу и перешли к фазам цветения и плодоношения практически одновременно с растениями более ранних сроков развития.
-
- ЦП 11, также описанной в синантропном сообществе, наибольшие значения CV отмечались для диаметра мужской корзинки и длины плода. Высокая степень изменчивости указанных параметров может являться следствием агротехнических мероприятий, в результате которых были обрезаны главные побеги растений. Это привело к формированию побегов второго порядка и, как следствие, выраженной морфометрической неоднородности генеративных структур.
Таким образом, при различных уровнях антропогенной и конкурентной нагрузок, вид демонстрирует свои адаптационные возможности через указанные параметры.
Между морфометрическими показателями во всех изученных ЦП наибольшее количество очень сильных положительных связей (r>0,8) было отмечено между длиной листа
- и двумя качественными признаками: шириной листа (6) и высотой побега (1) (табл. 2). Наибольшее число сильных корреляционных связей (r=0,71–0,8) проявлялось между высотой растения (1) и тремя качественными признаками: диаметром побега (2), шириной листа (6) и
89
Омельяненко Т. З., Багрикова Н. А.
![]()
длиной мужского верхушечного соцветия (7). Такое же число сильных связей было отмечено между длиной мужского верхушечного соцветия (7), высотой растения (1), длиной листа (5)
- его шириной (6). Таким образом, очень сильные и сильные корреляционные связи отмечаются внутри комплекса качественных параметров.
Таблица 2
Корреляционные связи морфометрических параметров всех ценопопуляций
Ambrosia artemisiifolia в 2021 году
| Параметры | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 1 | 0,00 | – | – | – | – | – | – | – | – | – | – | – |
| 2 | 0,73 | 0,00 | – | – | – | – | – | – | – | – | – | – |
| 3 | 0,66 | 0,66 | 0,00 | – | – | – | – | – | – | – | – | – |
| 4 | 0,56 | 0,72 | 0,53 | 0,00 | – | – | – | – | – | – | – | – |
| 5 | 0,81 | 0,58 | 0,58 | 0,41 | 0,00 | – | – | – | – | – | – | – |
| 6 | 0,78 | 0,63 | 0,59 | 0,46 | 0,93 | 0,00 | – | – | – | – | – | – |
| 7 | 0,77 | 0,59 | 0,51 | 0,35 | 0,75 | 0,73 | 0,00 | – | – | – | – | – |
| 8 | 0,56 | 0,39 | 0,44 | 0,25 | 0,54 | 0,5 | 0,63 | 0,00 | – | – | – | – |
| 9 | 0,25 | 0,43 | 0,3 | 0,31 | 0,27 | 0,34 | 0,38 | 0,13 | 0,00 | – | – | – |
| 10 | 0,47 | 0,63 | 0,36 | 0,7 | 0,32 | 0,36 | 0,32 | 0,13 | 0,2 | 0,00 | – | – |
| 11 | 0,24 | 0,25 | 0,21 | 0,15 | 0,23 | 0,26 | 0,26 | 0,22 | 0,4 | 0,06 | 0,00 | – |
| 12 | 0,12 | 0,24 | 0,14 | 0,2 | 0,09 | 0,14 | 0,17 | 0,04 | 0,29 | 0,13 | 0,5 | 0 |
Примечание к таблице. 1 – высота растения, см; 2 – диаметр побега, мм; 3 – количество ветвлений, шт.; 4 – количество листьев, шт.; 5 – длина листа, см; 6 – ширина листа, см; 7 – длина мужского верхушечного соцветия, см; 8 – количество мужских корзинок в соцветии, шт.; 9 – диаметр мужской (нижней в соцветии) корзинки, мм; 10 – количество плодов на одно растение, шт.;
- – длина плода, мм; 12 – ширина плода, мм.
Максимальное количество умеренных связей (r=0,61–0,7) свойственно для диаметра побега (2), количества ветвлений (3) и количества плодов (10) с другими вегетативными и генеративными признаками. Наибольшее число слабых связей (r=0,5–0,6) отмечалось для количества ветвлений (3) как с качественными, так и количественными вегетативными и генеративными параметрами.
Для признаков, коэффициент корреляции которых <0,5, выраженной взаимосвязи не отмечается. Например, для такого параметра, как диаметр мужской корзинки (9), не наблюдается ни одной корреляции, где r превышал бы 0,43. Для длины и ширины плода отмечается умеренная связь относительно друг друга, при этом от остальных параметров вида указанные признаки являются независимыми. Для семенной продуктивности отмечено две умеренных связи: с диаметром побега и количеством листьев. Со всеми остальными параметрами отмечается незначительная связь.
Изученные ценопопуляции статистически достоверно различаются в пространстве дискриминантных функций (p<0,01). На основании анализа квадратов расстояний Махаланобиса, установлено, что максимальные различия выявлены между ЦП 7 и ЦП 1–3, 6, 8, 10, 11, а также между ЦП 12 и ЦП 4, 5, 7, 9. При этом наибольшее расстояние отмечено непосредственно между ЦП 7 и ЦП 12 (61, 82). Таким образом, две указанные ценопопуляции являются наиболее фенотипически различными, как относительно друг друга, так и с остальными ценопопуляциями. ЦП 7 исследовалась в условиях выраженной нарушенности, в которой вид занимал одно из доминирующих положений в сообществе; ЦП 12, изученная при относительной сухости почв и высокой конкуренции со злаковыми и бобовыми видами, демонстрировала незначительную долю участия в сообществе и наименьшие средние значения, что подтверждает достоверность различий между указанными ценопопуляциями.
Минимальное расстояние отмечено между ЦП 1 и ЦП 2 (3,23), что отражает значительное сходство этих ценопопуляциий. Оба сообщества связаны с сегетальной растительностью, при этом ЦП 1 описана на периферии поля, ЦП 2 – на убранном поле зерновых. Отрастание побегов второго порядка после скашивания растений в ЦП 2 способствовало тому, что
90
Морфологическая изменчивость Ambrosia artemisiifolia в условиях Предгорного Крыма
![]()
некоторые параметры особей превышают таковые у растений, изученных в рудеральных, синантропных и синантропизированных сообществах. В частности, это прослеживается при рассмотрении таких параметров, как количество листьев и количество мужских корзинок в соцветии. Незначительное расстояние отмечено между ЦП 4 и ЦП 5 (3,83), при том, что условия местообитаний, время заноса вида и стадии сукцессионного процесса значительно различаются.
На основании кластерного анализа была построена дендрограмма, которая демонстрирует степени сходства и различия изученных ценопопуляций (рис. 2).
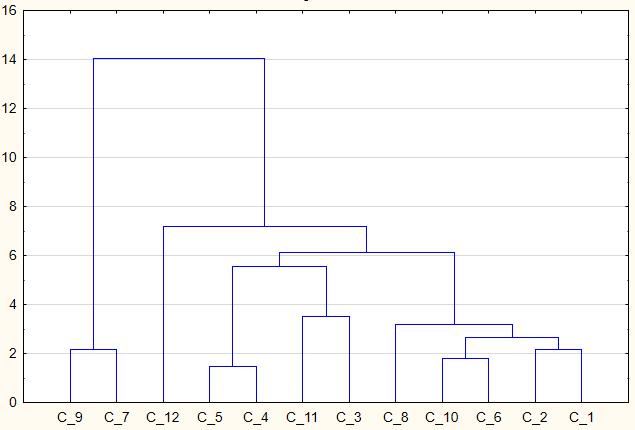
Рис. 2. Дендрограмма дифференциации ценопопуляций Ambrosia artemisiifolia
По оси абсцисс – № ценопопуляций, по оси ординат – Евклидово расстояние.
Согласно кластеризации методом k-средних ценопопуляции можно разделить на 4 кластера (табл. 3). При этом в первый кластер вошли ЦП 3, ЦП 4 и ЦП 5. Сходство ценопопуляций, в значительной мере, определено такими параметрами, как высота растения, количество ветвлений, длина и ширина листа, длина мужского соцветия и количество мужских корзинок в соцветии. В этих ценопопуляциях, описанных в синантропных, синантропизированных и рудеральных сообществах в Белогорском районе, при ОПП от 90 до 100 % на A. artemisiifolia приходилось от 50 до 75 % покрытия.
| Таблица 3 | ||||||||
| Элементы кластеров согласно кластеризации методом k-средних | ||||||||
| Расстояние | Расстояние | Расстояние | 4 | Расстояние | ||||
| 1 кластер | до центра | 2 кластер | до центра | 3 кластер | до центра | до центра | ||
| кластер | ||||||||
| кластера | кластера | кластера | кластера | |||||
| ЦП 3 | 0,78 | ЦП 1 | 0,48 | ЦП 12 | 0,00 | ЦП 7 | 0,56 | |
| ЦП 4 | 0,45 | ЦП 2 | 0,43 | ЦП 9 | 0,56 | |||
| ЦП 5 | 0,42 | ЦП 6 | 0,34 | |||||
| ЦП 8 | 0,61 | |||||||
| ЦП 10 | 0,43 | |||||||
| ЦП 11 | 0,65 | |||||||
| 91 | ||||||||
Омельяненко Т. З., Багрикова Н. А.
![]()
При детальном анализе значений морфометрических параметров, видно, что наибольшим сходством характеризуются ЦП 4 и ЦП 5, описанные в синантропизированных и рудеральных сообществах, что подтверждается наименьшим значением Евклидова расстояния между ними (1,39) (табл. 4).
Таблица 4
Евклидово расстояние между ценопопуляциями
| № ЦП | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 1011 | 12 |
| 1 | 0,00 | – | – | – | – | – | – | – | – | – | – | – |
| 2 | 2,08 | 0,00 | – | – | – | – | – | – | – | – | – | – |
| 3 | 3,71 | 4,29 | 0,00 | – | – | – | – | – | – | – | – | – |
| 4 | 2,55 | 2,76 | 4,19 | 0,00 | – | – | – | – | – | – | – | – |
| 5 | 2,46 | 3,41 | 4,02 | 1,39 | 0,00 | – | – | – | – | – | – | – |
| 6 | 2,16 | 2,33 | 4,23 | 3,61 | 3,52 | 0,00 | – | – | – | – | – | – |
| 7 | 6,50 | 7,88 | 6,15 | 6,99 | 6,16 | 7,37 | 0,0 | – | – | – | – | – |
| 8 | 3,55 | 2,19 | 5,47 | 3,45 | 4,13 | 2,70 | 8,7 | 0,00 | – | – | – | – |
| 9 | 4,64 | 6,29 | 4,40 | 5,32 | 4,59 | 5,84 | 3,9 | 7,26 | 0,00 | – | – | – |
| 10 | 2,28 | 2,43 | 4,55 | 3,33 | 3,52 | 1,74 | 7,5 | 2,65 | 5,83 | 0,00 | – | – |
| 11 | 2,87 | 3,33 | 3,45 | 3,64 | 3,53 | 2,50 | 6,4 | 3,63 | 4,49 | 3,16 | 0,00 | – |
| 12 | 5,57 | 4,74 | 7,72 | 5,12 | 5,97 | 5,09 | 10,5 | 3,37 | 8,76 | 4,31 | 5,64 | 0,0 |
Кластерный анализ позволил внести некоторые коррективы в трактование полученной дендрограммы. В частности, ЦП 11, которая на рисунке 2 представлена в общем кластере (ЦП 3, 4, 5 и 11), действительно имеет малое расстояние с ЦП 3, что выражается в близких значениях таких параметров, как высота растения, диаметр побега, количество листьев, длина листа, длина мужского верхушечного соцветия, диаметр корзинки, количество плодов и ширина плода. Обе ценопопуляции изучались в синантропных условиях.
Однако по данным кластеризации методом k-средних, ЦП 11 попадает во второй кластер. Анализ морфометрических показателей позволяет говорить, что для ЦП 11 отмечается наибольшее число близких значений с ЦП 1, ЦП 6 и ЦП 10, которые изучались в синантропных и рудеральных местообитаниях. Среди указанных ценопопуляций, наибольшим сходством характеризуются ЦП 10 и ЦП 6 (1,74) (табл. 4), что выражается в близких значениях большинства показателей, за исключением количества ветвлений и плодов. Обе ценопопуляции произрастают в рудеральных условиях в Симферопольском районе.
Несколько в меньшей мере сходство отмечено для ЦП 1 и ЦП 2 (2,08), изученных в синантропных и сегетальных сообществах в Белогорском районе. Достаточно близкими можно считать ЦП 2 и 8 (2,19), произрастающие в сегетальных и рудеральных местообитаниях. Помимо этого, сравнительно сходными являются ценопопуляции в рудеральных сообществах обоих районов – ЦП 8 и ЦП 10 (2,65). Ценопопуляции данного кластера в большей мере приурочены к рудеральным и синантропным условиям, в меньшей
– к сегетальным. Таким образом, ценопопуляции, объединенные во 2–й кластер, характерны для фитоценозов, испытывающих высокий уровень антропогенного пресса, в которых при ОПП от 55 до 100 %, на амброзию приходилось от 10 до 95 % покрытия.
На равном отдалении от центра 4-го кластера находится две образующие его ценопопуляции (ЦП 7 и ЦП 9), которые сходны по большинству параметров. Данные ценопопуляции отмечались в рудеральных и синантропных условиях Белогорского района, где на A. artemisiifolia приходилось от 50 до 90 %. Стоит отметить, что Евклидово расстояние между указанными ценопопуляциями составляет 3,9 и является наименьшим для этих
92
Морфологическая изменчивость Ambrosia artemisiifolia в условиях Предгорного Крыма
![]()
ценопопуляций. При этом для ЦП 7 со всеми остальными ценопопуляциями (кроме ЦП 3 и ЦП 9) отмечаются значительные расстояния (от 6,2 до 10, 5), что характеризует ЦП 7 как одну из наиболее обособленных ценопопуляций.
ЦП 12 занимает промежуточное место между ценопопуляциями, входящими во 2-ой и 4-ый кластеры и имеет наибольшее сходство с ценопопуляциями 2-го кластера. Однако относительно невысокие показатели габитуса и семенной продуктивности особей ЦП 12 не позволили отнести ее ни к одному кластеру. Наибольшее Евклидово расстояние отмечено между ЦП 12 и ЦП 3 (7,7), ЦП 7 (10,5), а также ЦП 9 (8,8), что характеризует их как наиболее отдаленные друг от друга. Наименьшее расстояние отмечено между ЦП 8 и ЦП 12 (3,37), что подтверждается некоторым сходством средних значений таких параметров, как высота растения, диаметр побега, количество листьев, длина и ширина листа, а также количество мужских корзинок в соцветии. Важно отметить, что проективное покрытие особей ценопопуляций также сходны – 15 и 20 %, соответственно. Таким образом, можно сделать вывод о том, что даже в разнообразных условиях произрастания, но при сходной степени конкуренции за ресурсы, может проявляться сходство растений по морфометрическим параметрам.
ВЫВОДЫ
В результате проведенных исследований установлено:
Вид обладает широким диапазоном значений морфометрических параметров вегетативных и генеративных органов, как в сравнительно сходных условиях, так и в достаточно отдаленных регионах Российской Федерации.
Наибольшими значениями морфометрических показателей характеризуется ценопопуляции, описанные в рудеральных и синантропных сообществах Белогорского района. Наименьшими значениями отличались особи, описанные в синантропизированном разнотравном сообществе, используемом для сенозаготовки.
Наибольший размах изменчивости и коэффициент вариации отмечены для количественных параметров вида. Наименьшей изменчивостью отличаются такие генеративные параметры, как длина и ширина плода, что говорит о выраженной стабильности этих показателей.
Наибольшее число сильных корреляционных связей было отмечено между высотой растения с диаметром побега, шириной листа и длиной мужского соцветия, что свидетельствует об их высокой согласованной изменчивости.
Наиболее фенотипически близкими согласно дискриминатному анализу можно считать ЦП 1 и 2, которые отмечались в синантропных и сегетальных сообществах Белогорского района. При этом, максимально отдаленными можно считать ЦП 7 и ЦП 12, изученных в рудеральных и синантропизированных условиях, соответственно.
По результатам кластерного анализа выделено 4 группы. Наибольшим сходством отличались ценопопуляции, которые отмечались преимущественно в условиях с выраженной антропогенной нагрузкой. Наименьшее Евклидово расстояние выявлено между ЦП 4 и 5, описанных в синантропизированных и рудеральных сообществах Белогорского района, что характеризует их как наиболее сходные.
Список литературы
Абрамова Л. М. Синантропизация растительности: закономерности и возможности управления процессом
(на примере Республики Башкортостан): автореф. дис. … докт. биол. наук: спец. 03.00.05 Ботаника, 03.00.16
Экология. – Пермь: Институт экологии Волжского бассейна РАН, 2014. – 45 с.
Багрикова Н. А., Скурлатова М. В. Материалы к «чёрной книге» флоры Крымского полуострова // Российский журнал биологических инвазий. – 2021. – Т. 14, № 2. – С. 16–31. DOI: 10.13140/RG.2.2.24139.72486
Беляева С. Н., Пирогова М. Е. Амброзия в Крыму: возможности элиминационной терапии // Вестник физиотерапии и курортологии. – 2020. – Т. 26, № 2. – С. 62–67.
Бурда Р. И. Антропогенная трансформация флоры / [Отв. ред. Кондратюк Е. Н]. – Киев: Наук. думка, 1991. – 168 с.
93
Омельяненко Т. З., Багрикова Н. А.
![]()
Голубев В. Н. Биологическая флора Крыма. – Ялта, НБС–ННЦ, 1996. – 126 с.
Голубев В. Н. Основы биоморфологии травянистых растений Центральной лесостепи. Часть I. Биоморфология подземных органов // Труды Центрально-Черноземного госзаповедника. – Воронеж: изд-во Воронежского ун-та, 1962. – Вып. 7. – С. 318–359.
Депозитарий живых систем «Ноев ковчег» [Электронный ресурс]. – Режим доступа: https://plant.depo.msu (просмотрено 11.02.2022)
Жалдак С. Н. Эколого-ценотические особенности Ambrosia artemisiifolia в условиях Предгорного Крыма // Экосистемы, их оптимизация и охрана. – 2011. – № 5 (24). – С. 66–70.
Злобин Ю. А. Принципы и методы ценотических популяций растений. – Казань: Казанский университет, 1989. – 146 с.
Кудрявцева Е. П., Базарова В. Б., Лящевская М.С., Мохова Л.М. Амброзия полыннолистная: современное распространение, структура сообществ и присутствие в голоценовых отложениях Приморского края (юг Дальнего Востока России) // Комаровские чтения. – 2018. – № 66. – С. 125–146. DOI: 10.25221/kl.66.5
Мамаев С. А. Основы проблемы исследования внутривидовой изменчивости растений // Флора и внутривидовая изменчивость растений Урала. – Свердловск, 1985. – С. 3–8.
Опанасенко Н. Е., Костенко И. В., Евтушенко А. П. Агроэкологические ресурсы и районирование степного
- предгорного Крыма под плодовые культуры. – Симферополь: ООО Издательство «Научный мир», 2015. – С. 70–
80.
Пархоменко В. М., Кашин А. С. Состояние ценопопуляций Hypericum perforatum L. в Саратовской области: Изменчивость морфометрических признаков и стратегия выживания // Растительные ресурсы. – 2011. – Т. 47, вып.
4. – С. 1–18.
Петрова С. Е. Онтогенез карантинных инвазионных сорняков Ambrosia artemisiifolia L. и A. trifida L. (Asteraceae) в Московской области // Российский журнал биологических инвазий. – 2019. – Т. 12, № 3. – С. 80–95.
Плантариум. Растения и лишайники России и сопредельных стран: открытый онлайн атлас и определитель растений. 2007–2022 [Электронный ресурс]. – Режим доступа: https://www.plantarium.ru (просмотрено 20.03.2022)
Тохтарь В. К. Дополнение к адвентивной флоре Юго-востока Украины// Украинский ботанический журнал. –
1996. – Т. 53, № 6. – С. 687–690.
Тохтарь В. К., Волобуева Ю. Е. Изменчивость семян Ambrosia artemisiifolia L. различного географического происхождения // Научные ведомости Белгородского государственного университета. Серия: Естественные науки. – 2012. – № 21 (140). – С. 19–22.
Тохтарь В. К., Петрик С. П. Одновременное появление адвентивных видов в разных регионах Украины // Украинский ботанический журнал. – 1993. – Т. 50, № 1. – С. 110–112.
Чадаева В. А., Шхагапсоева К. А., Цепкова Н. Л., Шхагапсоев С. Х. Мониторинг распространения Ambrosia artemisiifolia L. в луговых фитоценозах Кабардино-Балкарской Республики (Центральный Кавказ) // Российский журнал биологических инвазий. – 2018. – Т. 11, № 1. – С. 130–142.
Шварц Е. А. Сохранение биоразнообразия: сообщества и экосистемы. – М.: Т-во научных изданий КМК,
2004. – 112 с.
EPPO Global Database [Электронный ресурс]. – Режим доступа: https://gd.eppo.int (просмотрено 10.03.2022)
Global Biodiversity Information Facility [Электронный ресурс]. – Режим доступа: https://www.gbif.org (просмотрено 01.04.2022)
iNaturalist [Электронный ресурс]. – Режим доступа: https://www.inaturalist.org (просмотрено 20.03.2022)
Omelyanenko T. Z., Bagrikova N. A. Morphological variability of Ambrosia artemisiifolia in the Foothill Crimea // Ekosistemy. 2022. Iss. 30. P. 84–94.
Ambrosia artemisiifolia L. is a North American species and оne of the most widespread invasive plants in Russia, demonstrating high ecological plasticity. The paper presents the data about its distribution in the Republic of Crimea. The analysis of the variability of morphometric parameters of 12 cenopopulations studied in various ecotopes of the Foothill Crimea in 2021 was carried out. The highest values of morphometric indicators were registered in synanthropic and ruderal habitats, where communities are characterized by a significant level of anthropogenic load. The lowest average values were observed in synanthropized communities. Quantitative parameters have the greatest degree of variability. The qualitative parameters of the generative sphere were characterized by minimal variability. The correlation analysis revealed that the largest number of very strong and strong ties (r>0.71) was typical for qualitative parameters. Based on discriminant and cluster analyses, it was found that the greatest phenotypic similarity was specific for cenopopulations in synanthropized and ruderal habitats. Cluster analysis let the researchers combine 12 cenopopulations into four groups. The cenopopulation described in the synanthropized mixed-grass community was selected to a separate cluster. It differs from other cenopopulations by the lowest average values of morphometric parameters.
Key words: invasive plant, quarantine species, cenopopulations, variability, Foothill Crimea.
Поступила в редакцию 24.05.22
Принята к печати 23.06.22
94
