
ECOLOGICAL AND CENOTIC ANALYSIS OF PTERIDOFLORA SPECIES REGISTERED ON THE TERRITORY OF THE ABKHAZIAN FLORISTIC REGION OF THE CAUCASUS
JOURNAL: «EKOSISTEMY», Issue 25, 2021
Publication text (PDF): Download
UDK: 582.47(292.471):574.38
AUTHOR AND PUBLICATION INFORMATION
AUTHORS: Saltykov A. N., V.I. Vernadsky Crimean Federal University, Simferopol, Russia
TYPE: Article
DOI: https://doi.org/10.37279/2414-4738-2021-25-135-145
PAGES: from 135 to 145
STATUS: Published
LANGUAGE: Russian
KEYWORDS: Crimean pine, natural renewal, self-seeding, undergrowth, density, density, biogroup, spatial and age structure, age spectrum, ecological niche.
ABSTRACT (ENGLISH):
Studies of the processes of natural renewal of Crimean pine, carried out during 2018–2019, on the mountain ranges of the Yalta Mountain Forest Reserve allowed us to establish a number of the following features. The surge of renewal is transient in time and the basis of the coenopopulations of undergrowth at the object of research is made up of plants that appeared in the first two post-fire years. Individuals that appeared in subsequent years occupy vacant places in the conditions of existing renewal niches, complementing the structure of the age spectra of pine undergrowth, emphasizing the asymmetry of the spectral distribution series. On the one hand, the spatial structure of undergrowth coenopopulations is a representation of the “matrix” of forest fire, on the other hand, it is determined by the structure of pine trees’ fruiting. The monotonous decrease in the density of the population field of undergrowth, due to the remoteness of the walls of the mother plantings and the specifics of seed dispersal, is supplemented by the alternation of undergrowth biogroups of increased density. The presence of areas of increased density against the background of constantly decreasing density of plants with a distance from the walls of the mother plant is predetermined by the influence of testes and seed curtains preserved during the fire. At each specific point in the renewal space, the structure of the process or its shape will be determined by the influence of external environmental and internal endogenous factors due to the biogroup effect of plant groupings. Polyvariance of the spatio-age structure of the undergrowth in the gorelnik space is the result of autoregulation of the cenopopulation structure in accordance with the capacity of the existing renewal niches.
ВВЕДЕНИЕ
Сохранение биологического разнообразия и устойчивости лесных экосистем является одним из актуальных направлений в исследованиях отечественного лесоводства (Рожков, 1989; Демаков, 2000; Восточно-европейские леса…, 2004; Бех, 2015; Коротков, 2015; Коба, 2016; Салтыков, 2019).
Бурное развитие популяционной экологии на фоне огромного количества накопленного материала, позволило углубить и даже изменить устоявшееся мнение об особенностях процессов естественного возобновления, устойчивости и продуктивности лесных экосистем (Злобин, 1976; Рифлекс, 1979; Одум, 1986; Бигон, 1989; Рожков, 1989; Демаков, 2000; Бех, 2015).
Так, в рамках популяционно-онтогенетического направления, биоценотический покров рассматривается как комплекс взаимодействующих популяций, а, учитывая эдафическую однородность биоценоза, ценопопуляций растений. Сформулировано понятие ценопопуляции подроста и разработаны методические подходы оценки качественного её состояния (Злобин, 1976). Исследованиями установлено, что ценопопуляции в границах лесного биоценоза свойственна различная степень гетерогенности в структурном, динамическом и функциональном аспектах (Грейг-Смит, 1967; Восточно-европейские леса…., 2004; Коба, 2016, 2017; Сприцын, 2017; Малиновских, 2017; Салтыков, 2019; Санников, 2019).
Переход от статичных моделей количественной оценки процессов естественного возобновления к основам оценки качественной структуры ценопопуляции позволил выявить динамику и направленность процесса (Злобин, 1976; Салтыков, 2019; Санников, 2019).
Очевидно, что одним из перспективных направлений сохранения биологического разнообразия и устойчивости является восстановление коренных древостоев, а, следовательно, использование естественного потенциала лесных экосистем (Пятницкий, 1959; Рожков, 1989; Демаков, 2000; Грязькин, 2001; Ерохин, 2011; Коротков, 2015; Бех, 2015; Коба, 2016, 2017, 2018; Малиновский, 2017; Спринцын, 2017; Грязькин, 2019; Салтыков, 2019; Санников, 2019).
Структурно-функциональные особенности процесса возобновления, должны служить основой для разработки лесоводственных мероприятий, позволяющих реализовать в практике хозяйствования принципы устойчивого ведения лесного хозяйства и устойчивого лесоуправления (Злобин, 1976; Грязькин, 2001; Салтыков, 2019; Санников, 2019).
В силу ряда объективных причин, до настоящего времени отсутствуют сведения о закономерностях формирования популяционных мозаик, причинах и следствиях поливариантности структуры ценопопуляций, механизмах меж- и внутрипопуляционного взаимодействия и влияния растений и так далее (Восточно-европейские леса…, 2004).
Целью наших исследований являлось изучение особенностей пространственно-возрастной структуры подроста сосны крымской (Pinus рallasiana D. Don) и процессов авторегуляции указанной структуры в соответствии с ёмкостью существующих ниш возобновления.
МАТЕРИАЛ И МЕТОДЫ
В течение 2018–2019 годов исследования проводились на территории горельника Алупкинского научно-исследовательского природоохранного отделения Ялтинского горно-лесного заповедника, который возник в результате пожара в августе 2007 года. Лесным пожаром было уничтожено около 1000 га спелых и перестойных коренных древостоев сосны крымской. В 2008–2009 годах выполнена сплошная санитарная рубка повреждённого древостоя. Уцелевшие во время пожара одиноко стоящие деревья и куртины были оставлены в качестве семенников. Фрагменты насаждений по внешним контурам горельника, пройдённые беглым низовым пожаром, и не повреждённые огнём, также не вовлекались в рубку. Всплеск естественного возобновления пришёлся на последующие два года.
Результаты рекогносцировочных исследований показали, что подрост сосны (P. рallasiana) на площади горельника присутствует повсеместно, но пространственная структура и жизненное его состояние неоднородны. Специфика размещения подроста в пространстве исследуемого объекта и его качественное состояние были приняты нами во внимание при формировании сети пробных площадей по вариантам опыта. Так, например, в первом случае серия пробных площадей была заложена непосредственно под пологом древостоя. Во втором варианте серия пробных площадей размещена в 50 метровой зоне прилегающей к стене материнского насаждения, в границах которой прослеживается её влияние на рост и развитие подроста сосны. Следующие варианты опыта с целью исключения такого влияния размещены на 100 метровом удалении от стены леса и за его пределами.
При выполнении рекогносцировочных исследований под пологом и за границами лесных насаждений нами была заложена серия учётных площадок размером 1×1 (м2) и 1×2 (м2) (Грейг-Смит П., 1967). Общее число учётных площадок составило 100 шт. После выявления общих закономерностей формирования подроста сосны крымской в границах существующих ценопопуляций подроста нами были заложены пробные площади размером 10×10 (м2) согласно методике С. С. Пятницкого (1959). В каждом конкретном варианте опыта предусмотрена четырёх- или же восьмикратная повторность. Под кронами материнских насаждений наряду с указанной методикой заложена серия учётных пробных площадок размером 5×5 (м2) в количестве 10 шт. Общее число площадок, заложенных по методике С. С. Пятницкого составило 72 шт. На указанных пробных площадях для подроста сосны устанавливались следующие показатели: замерялась высота (см) и количество растений на пробе. Для тридцати случайно отобранных растений устанавливали: диаметр на шейке корня и высоте груди (см), возраст (лет), верхушечный прирост за последний год (см), ширину кроны в противоположных направлениях (см), состояние растения. Полученные данные обработаны общепринятыми методами математической статистики. Всего при выполнении исследований заложено 182 учётные пробные площади и выполнена биометрическая оценка более 3 тыс. растений. Полученные данные положены в основу анализа особенностей пространственно-возрастной структуры подроста сосны крымской и процессов авторегуляции структуры ценопопуляции.
В рамках выполненных нами исследований используется понятие ценопопуляции подроста сосны, предложенное Ю.А. Злобиным, который писал: «При возобновлении на вырубках и гарях ценопопуляция подроста древесной породы соответствует ценопопуляции данного вида в её полном объёме. В случае возобновления под пологом материнской породы совокупность особей подроста является лишь частью ценопопуляции вида. В связи с экологической самостоятельностью подроста такую субценопопуляцию подроста допустимо рассматривать отдельно» (Злобин, 1976).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Исследованиями 2018 года установлено, что подрост P. рallasiana распространён практически по всей площади горельника, но его состояние, особенности роста и развития заметно варьируют. Под пологим материнским древостоем подрост отстаёт в росте, состояние растений угнетённое. Влияние материнского насаждения на подрост прослеживается и в непосредственной близости к стенам леса, с удалением от которых можно наблюдать снижение плотности биогрупп и синхронное увеличение биометрических показателей растений: средней высоты, среднего диаметра и размеров кроны. Варьирование плотности растений в границах существующих ценопопуляций достаточно хорошо различимо, зависимость между густотой и биометрическими показателями растений выражена менее отчётливо. Итоги дисперсионного анализа отражены в таблицах 1 и 2.
Таблица 1
Сравнительная оценка подроста сосны крымской по высоте, выполненная на основании результатов дисперсионного анализа
| Вариант | Подрост на удалении от стены насаждения | Средняя высота, см | Изменение высоты, см |
| 1 | на расстоянии до 50 м | 137,2 | – |
| 2 | на расстоянии от 50 до 100 м | 166,6 | 29,4 |
| 3 | на расстоянии свыше 100 м | 175,1 | 37,9 |
| Наименьшая существенная разность (НСР) 0,95 | – | 25,2 | |
Фактическая разница средних высот растений во всех вариантах опыта превышает наименьшую существенную разность по рассматриваемому показателю при уровне вероятности 0,95. Максимальная средняя высота растений зафиксирована на значительном удалении, минимальная в непосредственной близости от стены насаждения (табл. 1). Таким образом, с удалением от стены материнского насаждения на фоне постепенного снижения густоты растений можно наблюдать увеличение комплекса биометрических показателей и, прежде всего, средней высоты подроста. В зонах, где влияние леса сведено к минимуму, успешность роста сосны, вероятно, сопряжена с плотностью биогрупп.
С целью проверки зависимости между плотностью стояния растений и комплексом биометрических показателей, нами также была заложена сеть пробных площадей. Варианты опыта изначально разделены на две составляющие. В первом случае густота растений превышает 10 тыс. шт./га, во втором она в 1,5–2 раза меньше указанной величины. Удаление от стены материнского насаждения во всех случаях составило более 100 м. Результаты дисперсионного анализа отражены в таблице 2.
Таблица 2
Сравнительная оценка подроста сосны крымской по высоте, выполненная на основании результатов дисперсионного анализа
| Вариант | Количество подроста тыс. шт./га. | Средняя высота, см | Изменение высоты, см |
| 1 | до 13,5 | 128,1 | – |
| 2 | до 13,0 | 131,8 | 30,6 |
| 3 | до 7 (6,8) | 158,7 | 26,9 |
| Наименьшая существенная разность (НСР) 0,95 | – | 15,7 | |
| Наименьшая существенная разность (НСР) 0,99 | – | 24,2 | |
Примечание к таблице. Удаление подроста сосны крымской от стены материнского насаждения составляет более 100 м.
Фактическая разность средних высот превышает наименьшую существенную разность по средней высоте биогрупп при уровне вероятности 0,95 и 0,99 в двух вариантах опыта из трёх. То есть правомерно утверждение о наличии статистически достоверной разницы в результатах исследования.
Известно, что в биогруппах подроста с нарастанием конкуренции неизбежна дифференциация, расслоение особей на отставшие в росте, угнетённые и успешно произрастающие (Гончар, 1957; Малиновских, 2017; Салтыков, 2019, Санников, 2019). Последующее самоизреживание группы, как следствие авторегуляции структуры ценопопуляции сопровождается сравнительно успешным ростом сосны, сохранившей за собой лидирующее положение.
Процессы динамики структуры ценопопуляций подроста находят своё подтверждение в конкретных биометрических показателях растений на опытных объектах (табл. 1, 2, 4–6) и являются отражением установленных закономерностей пространственной структуры ценопопуляции. Например, отчётливо выражено убывание численности подроста сосны с удалением от стен материнских насаждений (рис. 1). Синхронно изменению густоты варьируют и биометрические показатели подроста.
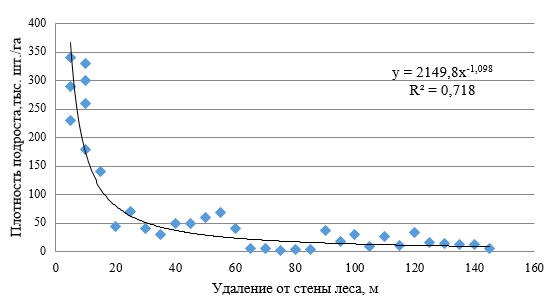
Рис. 1. Зависимость между густотой (плотностью) подроста сосны крымской и его удалением от стены прилегающего материнского насаждения
Очевидная разница в комплексе биометрических показателей растений, приуроченных к подкроновому пространству, расположенных в непосредственной близости к стенам леса и удалённых на значительное расстояние от материнского насаждения, является основанием для выдвижения гипотезы о том, что причиной гетерогенности структуры ценопопуляций является их возрастная структура. Процесс плодоношения и последующей реализации репродуктивного потенциала насаждений в категорию самосева и подроста по целому ряду причин мог быть растянут во времени. Можно предположить, что увеличение средней высоты, среднего диаметра, кроны подроста синхронно увеличению его возраста. В тоже время, варьирование плотности существующих ценопопуляций подроста и биометрических показателей растений в равной мере могло быть вызвано влиянием материнского насаждения посредством формирования экологических условий, в той или иной мере лимитирующих рост и развитие растений. Поливариантность реализации процесса возобновления в пространстве горельника в каждом конкретном случае является следствием подобных закономерностей.
С целью выявления причин, повлёкших варьирование структуры ценопопуляций, в 2019 году исследования были продолжены. Итоги изучения возрастной структуры подроста сосны позволили установить, что доминирующая его часть на открытых площадях горельника достигла 10–11 летнего возраста, а, следовательно, его появление следует датировать 2008–2009 годами (рис. 2, 3).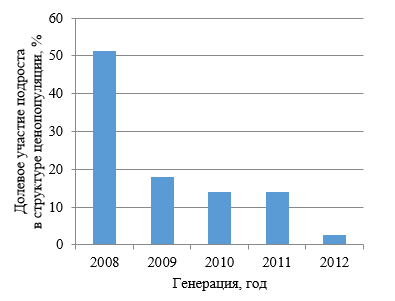
Рис. 2. Возрастной спектр подроста сосны крымской за пределами влияния материнского насаждения
Кроме 10 и 11 летних особей в границах возрастных спектров подроста присутствуют растения, которые появились на третий, четвёртый и даже пятый год после пожара. Указанная категория растений, как правило, располагается по периферии или же занимает подчиненное место в границах сложившихся биогрупп, долевое их участие в количественном отношении незначительно. Заполняя свободные места в нишах возобновления, указанная категория растений способствует формированию уплотнённых группировок с характерной для них микросредой и успешным ростом сосны.
Под пологом материнских насаждений состояние подроста и комплекс биометрических характеристик растений заметно отличается. Так, например, средний возраст особей здесь сравнительно меньше, чем на открытых пространствах горельника (табл. 3).
В границах возрастного спектра, кроме десяти и одиннадцатилетних, присутствуют особи, которые появились через шесть, семь и восемь лет после пожара. Обращает на себя внимание доминирование особей, появившихся в 2010 и 2012 годах (рис. 3). Вероятно, снижение среднего возраста подроста под пологом материнских насаждений (6,9±0,06 лет)
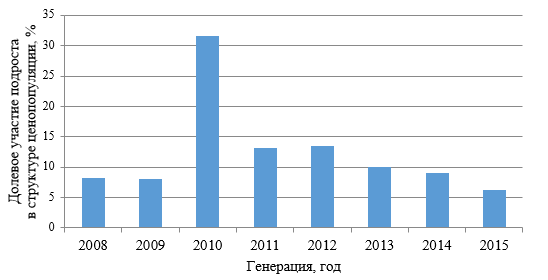
Рис. 3. Возрастной спектр подроста сосны крымской под пологом насаждений
Таблица 3
Варьирование среднего значения возраста растений подроста сосны крымской в зависимости удаления от стены материнского насаждения
| Показатель | Удаление от стены материнского насаждения, м | ||||
| под пологом | 10–50 | 51–100 | >100 | ||
| Возраст, x̅±S x̅ | 6,9±0,06 | 9,3±0,08 | 9,7±0,17 | 9,9±0,20 | 10,1±0,14 |
| Эксцесс | –0,65 | –0,19 | –0,84 | –0,44 | –0,51 |
| Асимметрия | –0,32 | –0,92 | 0,38 | –0,87 | –0,90 |
| Точность опыта | 0,8 | 0,8 | 1,7 | 2 | 1,4 |
| Варьирование | 28,3 | 8,8 | 10,4 | 12,2 | 12 |
обусловлено двумя встречными процессами: отпадом подроста, появившегося в первые годы после пожара и наличием растений новых генераций. Причина отпада старших по возрасту растений очевидна. Жизнеспособность подроста P. рallasiana под пологом материнского насаждения невысокая, и по мере исчерпания экологической ниши, подрост сосны уступает своё место следующим поколениям или замещается растениями – экологическими аналогами, типичными для данных условий местопроизрастания. С определённой степенью условности можно сказать, что волна возобновления в данных условиях скоротечна во времени и динамична в пространстве, что согласуется с мнением исследователей (Коба, 2016, 2017; Санников, 2019).
Сравнение структуры возрастного спектра подроста на открытых пространствах и под пологом материнских насаждений (рис. 2, 3), а также сведения о среднем возрасте растений позволяют выдвинуть предположение о том, что «стартовые» позиции процесса естественного возобновления под пологом насаждений и на открытых пространствах горельника были близкими. Однако со временем условия экологической ниши, в которой развивался подрост, отразились на его биометрических характеристиках (табл. 2, 3, 4). Так, например, средняя высота подроста под пологом материнского насаждения при среднем возрасте от 7,3±0,12 до 8,1±0,16 лет колеблется в пределах 21,5±1,09 – 27,4±1,09 см, прирост верхушечного побега варьирует от 3,6±0,11 до 4,8±0,17 см, при этом минимальное его значение составляет 2,4±0,21 см (табл. 2). Густота растений составляет от 16 до 63,2 тыс. шт./га. На первый взгляд, это очень большая численность растений. Однако число растений в количестве 1–2 шт./м2 или даже 6–7 шт./м2, при их средней высоте от 13,1±0,80 до 27,4±1,09 и среднем приросте не превышающем 3,8±0,19 см (табл. 4) позволяет поставить под
Таблица 4
Биометрическая оценка подроста сосны крымской под пологом лесных насаждений
| № | Диаметр шейки корня, см | Высота растения, см | Возраст, лет | Прирост, см | Плотность | ||
| текущий | средний | шт./м2 | тыс. шт./га | ||||
| 1 | 0,4±0,02 | 13,1±0,8 | 3,8±0,19 | 2,4±0,21 | 3,8±0,19 | 1–2 | 16,0 |
| 2 | 0,5±0,04 | 22,9±1,45 | 7,3±0,18 | 3,8±0,22 | 3,1±0,19 | 3–4 | 30,4 |
| 3 | 0,7±0,05 | 21,5±1,09 | 8,1±0,16 | 3,6±0,11 | 2,7±0,14 | 3–4 | 38,8 |
| 4 | 0,6±0,06 | 21,4±0,75 | 7,0±0,12 | 3,5±0,13 | 2,8±0,10 | 4–5 | 42,8 |
| 5 | 0,7±0,06 | 19,3±0,74 | 6,8±0,14 | 3,4±0,15 | 2,9±0,12 | 4–5 | 44,4 |
| 6 | 0,7±0,03 | 27,4±1,09 | 7,3±0,12 | 4,8±0,17 | 3,9±0,18 | 5–6 | 58,0 |
| 7 | 0,6±0,04 | 20,6±0,60 | 7,4±0,14 | 3,8±0,15 | 2,8±0,09 | 5–6 | 59,2 |
| 8 | 0,6±0,03 | 15,6±0,63 | 5,7±0,15 | 2,6±0,12 | 2,7±0,11 | 6–7 | 63,2 |
Примечание к таблице. По каждому из показателей приведены средние значения ± ошибка средней. В первой колонке приведены номера пробных площадок.
сомнение успешность процесса естественного возобновления сосны крымской под пологом материнского насаждения.
Присутствие 10- и 11-летних особей в границах возрастных спектров, хорошо выраженная временная граница начала спектральной характеристики растений по возрасту (2008 г.), сходство шлейфа сопутствующих возрастных генераций подроста позволяет утверждать, что подрост сосны крымской под пологом и за пределами влияния материнских насаждений по своему происхождению может быть отнесен к единому пространству возобновления. Однако по мере исчерпания ёмкости экологической ниши затухание всплеска возобновления под пологом материнских насаждений закономерно. В результате подрост P. рallasiana под пологом насаждений представляет собой очень динамичный и скоротечный во времени фрагмент ценопопуляции. Снижение среднего возраста подроста в этих условиях позволяет объяснить направление и динамику процесса. Ожидаемое количественное и пространственное перераспределение ценопопуляции происходит вследствие несовпадения ёмкости экологической ниши и биоэкологических свойств растений на определенном этапе их развития (Злобин, 1976; Коба, 2016, 2017; Малиновский, 2017; Сприцын, 2017; Салтыков, 2019; Санников, 2019). Подрост сосны в сравнительно короткий временной период уступает своё место для следующих поколений, а зоны краевого эффекта смещаются на периферию открытых пространств горельника. Так, например, максимальные показатели плотности ценопопуляции подроста P. рallasiana были отмечены нами в пределах 50-метровой зоны прилегающей стены материнского насаждения, где количество подроста достигает 60–70 тыс. шт./га и более. На площадках, размещенных на 10 метровом удалении от стены леса нами зафиксированы биогруппы густотой от 23 до 40 растений на 1 м². Среднее количество растений в непосредственной близости к стенам материнских насаждений составило 30,2±1,63 шт./м2. С удалением от стены насаждения на расстояние свыше 100 метров количество подроста уменьшается до 10–20 тыс. шт./га. Таким образом, одна из характерных черт пространственной структуры ценопопуляций подроста на открытых пространствах горельника – резкое увеличение численности подроста вблизи стен материнских насаждений и снижение плотности биогрупп с удалением к центральной части горельника (рис. 1).
В тоже время на фоне постепенного изменения численности подроста можно наблюдать сравнительно плотные растительные группировки, появление которых обусловлено влиянием семенных деревьев и куртин, оставшихся после разработки горельника. Варьирование плотности растительных группировок сопровождается синхронным изменением биометрических характеристик подроста сосны (табл. 3). Так, только в пределах 25-метровой зоны средняя высота подроста сосны увеличивается в два и более раза, достигая 190,6±14,47 см, средний прирост верхушечной оси составляет 20,6±1,48 см. (табл. 3). На пятидесятиметровом удалении максимальная величина средней высоты подроста сосны составляет от 190,0±14,91 до 195,9±10,96 см. (табл. 3). В этих же условиях заметно возрастает текущий прирост верхушечной оси, достигая 22,8±3,64 см, превышая отметку среднего прироста, что позволяет отнести рассматриваемые фрагменты ценопопуляции подроста к процветающему типу, то есть весьма успешно развивающейся категории растений. Густота стояния растений на указанном удалении от стены материнского насаждения колеблется от 40 до 60 тыс. шт./га (табл. 5).
Таблица 5
Биометрическая оценка подроста сосны крымской, расположенного на расстоянии до 50 м от стены материнского насаждения
| № | Диаметр шейки корня, см | Высота растения, см | Возраст, лет | Прирост, см | Плотность | ||
| текущий | средний | шт./м2 | тыс. шт./га | ||||
| На удалении до 25 м от стены материнского насаждения | |||||||
| 1 | 2,2±0,09 | 94,1±3,99 | 9,3±0,17 | 12,5±0,17 | 10,3±0,39 | 18 | 180 |
| 2 | 2,2±0,17 | 132,8±5,37 | 9,2±0,21 | 15,9±0,10 | 14,8±0,55 | 14 | 140 |
| 3 | 3,1±0,34 | 189,7±13,35 | 9,2±0,22 | 16,6±1,98 | 20,6±1,48 | 4 | 45 |
| 4 | 3,4±0,29 | 190,6±14,47 | 9,8±0,21 | 18,0±1,06 | 19,6±1,55 | 7 | 70 |
| На удалении до 50 м от стены материнского насаждения | |||||||
| 1 | 2,7±0,27 | 150,1±13,22 | 8,7±0,29 | 18,2±3,23 | 17,1±1,15 | 7–8 | 75 |
| 2 | 3,2±0,31 | 190,0±14,91 | 8,8±0,31 | 20,3±1,28 | 21,6±1,44 | 3–4 | 40 |
| 3 | 3,5±0,75 | 195,9±10,96 | 9,3±0,18 | 21,3±1,62 | 21,1±1,03 | 5–6 | 60 |
| 4 | 3,8±0,75 | 177,6±23,35 | 9,0±0,57 | 22,8±3,64 | 19,8±2,34 | 3–4 | 40 |
Примечание к таблице. По каждому из показателей приведены средние значения ± ошибка средней. В первой колонке приведены номера пробных площадок.
В границах плотных биогрупп средние: высота, диаметр растений, размер кроны заметно ниже, чем в разреженных. Динамика рассматриваемых средних показателей растений является основанием для предположения о том, что в успешный рост сосны будет наблюдаться при определённой густоте их размещения. Снижение или увеличение комплекса биометрических характеристик подроста P. рallasiana в границах сформировавшихся ценопопуляций является ответной реакцией подроста на изменение экологических условий, обусловленных плотностью популяционного поля. Таким образом, в рамках процесса естественного возобновления отчётливо проявляется «эффект биогруппы». Наличие относительно плотных биогрупп на начальных этапах становления ценопопуляции определяет успешный рост особей, исключая конкуренцию со стороны растений экологических аналогов (Гончар, 1957; Малиновский, 2017; Салтыков, 2019; Санников, 2019. На последующих этапах роста и развития этот показатель будет изменяться. Подтверждением являются результаты биометрической оценки подроста, расположенного в границах 100-метровой зоны от стен материнского насаждения (табл. 6).
Характерно, что в границах указанной зоны заметно снижается густота растений на единице площади. Варьирование средней высоты подроста незначительно и составляет от 158,0±6,28 до 197,1±9,07 см. Доминирует 10-летний подрост, средний возраст растений равен 9,7±0,17 лет. В целом следует сказать, что в условиях, исключающих влияние материнского насаждения, сохраняется биогрупповой характер размещения растений в пространстве, подрост сосны отличается хорошим ростом и имеет перспективу дальнейшего роста и развития. Неизменным остаётся и постоянство чередования разных по плотности биогрупп: уплотнённых и редких. Доля не занятых подростом сосны площадей незначительна. В большинстве своём такие прогалины, поляны, «окна», где отсутствует подрост сосны крымской, составляет около 30–35 % от общей площади горельника. Зачастую подрост сосны
Таблица 6
Биометрическая оценка высоты подроста сосны крымской, расположенного на расстоянии 50–100 м от стен материнских насаждений
| Показатель | Высота подроста, см
Номер варианта и среднее значение высоты (x̅±S x̅) по вариантам |
||||
| 1 | 2 | 3 | 4 | 1–4 | |
| 158,0±6,28 | 183,5±8,60 | 197,1±9,07 | 161,7±8,46 | 171,8±4,12 | |
| Эксцесс | –0,49 | 0,34 | –0,59 | –0,27 | 0,01 |
| Асимметрия | 0,14 | 0,71 | 0,01 | 0,74 | 0,55 |
| Точность опыта, % | 3,90 | 4,70 | 4,60 | 5,20 | 2,00 |
| Варьирование, % | 31,00 | 33,50 | 24,30 | 35,50 | 32,70 |
| Плотность, тыс. шт./га | 6,10 | 5,10 | 2,80 | 4,60 | 4,70 |
отсутствует на местах хранения древесины или погрузочных площадках, сети технологических элементов лесосек, которые были использованы при проведении сплошных санитарных рубок. Очевидно, что такие разрывы в границах формируемого насаждения со временем будут заняты последующими генерациями P. рallasiana, что в определённой мере окажет влияние на особенности пространственной структуры формируемых коренных древостоев.
Таким образом, в результате прохождения лесного пожара и последовавшего за ним всплеска возобновления на территории горельника наблюдается формирование единого структурно-функционального поля возобновления. С определённой степенью условности такое поле можно разделить на три взаимосвязанные составные части. Первая – это зона с подростом сосны, сосредоточенным в подкроновом пространстве насаждений. Это незначительная по площади затронутая низовым пожаром часть насаждений по внешним границам горельника. Подрост сосны в этой зоне, как правило, угнетён и не жизнеспособен. Вторая зона с подростом сосны, расположенным в непосредственной близости к стенам материнского насаждения. Характерным признаком такой зоны является чрезвычайно высокая плотность и динамичное состояние подроста. Третья – зона с подростом сосны, удалённым на значительное расстояние от стен материнских насаждений. Плотность подроста незначительна, жизненное состояние ценопопуляций подроста большей частью, процветающее. В границах второй и третьей зоны на фоне закономерно убывающей численности подроста с удалением от стены материнского насаждения можно наблюдать чередование незначительных по плотности и уплотнённых растительных группировок, кластерных зон, оказывающих заметное влияние на структурно-функциональное единство процесса возобновления.
ЗАКЛЮЧЕНИЕ
Исследования структурно-функциональных особенностей естественного возобновления сосны крымской (P. рallasiana) на территории горельника (2007 года) Ялтинского горно-лесного заповедника позволили сделать вывод о том, что всплеск возобновления достаточно скоротечен во времени и в данном случае он пришёлся на 2008–2009 годы.
В настоящее время большая часть горельника занята жизнеспособным подростом сосны крымской. Доля не занятых подростом сосны площадей здесь незначительна. Прогалины и поляны, где отсутствует подрост сосны крымской, занимают немногим более третьей части площади объекта исследования.
Максимальная густота растений, достигающая 50–70 тыс. шт./га и более, наблюдается в непосредственной близости к стенам материнского насаждения. С удалением от стены насаждения на расстояние до 100 метров количество подроста уменьшается до 10–20 тыс. шт./га. На фоне снижения плотности биогрупп наблюдается синхронное увеличение биометрических показателей растений: средней высоты, среднего диаметра и размеров кроны. На открытых пространствах горельника подрост сосны большей частью, жизнеспособный. Варьирование плотности растительных группировок обусловило гетерогенность пространственной структуры ценопопуляций подроста сосны крымской.
Под пологом материнского насаждения жизнеспособность подроста сосны невысокая, и по мере исчерпания экологической ниши, наблюдаются процессы экологического замещения. Можно утверждать, что волна возобновления в данных условиях скоротечна во времени и динамична в пространстве, что в полной мере согласуется с мнением других исследователей.
Единое структурно-функциональное поле возобновления условно можно разделить на три взаимосвязанные составные части. Первая – это зона с подростом сосны, сосредоточенным в подкроновом пространстве насаждений. Вторая зона с подростом сосны, расположенным в непосредственной близости к стенам материнского насаждения. Третья – зона с подростом сосны, удалённым на значительное расстояние от стен материнских насаждений. В границах указанных зон состояние и особенности роста и развития растений заметно отличаются.
Поливариантность реализации процесса возобновления в пространстве горельника в каждом конкретном случае является результатом адаптации подроста сосны крымской и авторегуляции структуры ценопопуляции подроста в соответствии с ёмкостью сложившихся ниш возобновления, их соответствия биоэкологическим свойствам подроста на определённом этапе его роста и развития.
REFERENCES
Бех И. А., Данченко А. М. Проблема устойчивости в лесоведении // Вестник Томского государственного университета. — 2015. — № 295. — С. 215-219.
Бигон М., Харпер Дж., Таусенд К. Экология. Особи, популяции и сообщества. В 2-х томах / [Ред. А. М. Гиляров]. — Москва: Мир, 1989. — Т. 1. — 667 с., Т. 2. — 447 с.
Восточноевропейские леса: история в голоцене и современность. В 2 кн. / [Отв. ред. О. В. Смирнова]. – М.: Наука, 2004. Кн. 1. – 479 c.; Кн. 2. – 575 с.
Гончар М. Т. О влиянии группового произрастания соснового подроста на условия микросреды // Записки Харьковского сельскохозяйственного института. – 1957. – Т. XVI. – С. 135–150.
Грейг-Смит П. Количественная экология растений / [Ред. Т. А. Работнов и А. А. Уранов]. — Москва: Мир, 1967. — 359 с.
Грязькин А. В. Возобновительный потенциал таёжных лесов (на примере ельников Северо-Запада России). — СПб.: Санкт-Петербуржская государственная лесотехническая академия (СПбЛТА), 2001. — 188 с.
Демаков Ю. П. Динамика устойчивости лесных экосистем: (Методологические и методические аспекты). — Йошкар-Ола: Периодика Марий Эл, 2000. — 416 с.
Ерохин А. В. Естественное возобновление в культурах сосны // Актуальные проблемы лесного комплекса. — 2011. — Вып. 30. — С. 13 — 15.
Грязькин А. В., Беляева Н. В., Шахов А. Г., Нгуен Ван Зинь Естественная смена ели сосной на участках лесных культур // Лесотехнический журнал. — 2019. — Т. 9, № 1 (33). — С. 54-60.
Злобин Ю. А. Оценка качества ценопопуляций подроста древесных пород // Лесоведение. – 1976. – № 6. – С. 72–79.
Коба В. П., Жигалова Т. П. Возобновление сосны Палласа на горельниках в горном Крыму // Лесоведение. — 2016. – № 4. – С. 270–278.
Коба В. П. Особенности восстановления древостоев сосны крымской в постпирогенный период // Экосистемы. — 2017. – Вып. 11. – С. 10–13.
Коба В. П., Жигалова Т. П. Экспериментальная оценка постпирогенной жизнеспособности семян сосны Палласа // Лесоведение. — 2018. – № 6. – С. 417–425.
Коротков С. А. Теоретические проблемы устойчивости леса // Лесной вестник. — 2015. — № 4. — С. 26-32.
Малиновских А. А. Динамика естественного возобновления сосны обыкновенной на гари 1977 года в Сростинском Бору // Вестник алтайского аграрного университета. — 2017. — № 5 (151).- С. 76-82.
Одум Ю. Экология. В 2-х томах / [Ред. В. Е. Соколова]. — Москва: Мир, 1986. — Т. 1. — 328 с., Т. 2. — 376 с.
Пятницкий С. С. Методика исследований естественного семенного возобновления в лесах левобережной Лесостепи Украины. – Харьков, 1959. – 38 с.
Риклефс Р. Основы общей экологии / [Ред. Н. Н. Карташев]. – Москва: Мир, 1979. – 424 с.
Рожков А. А., Козак В. Т. Устойчивость лесов. — Москва: Мир, 1989. — 238 с.
Салтыков А. Н. Структурно-функциональные особенности естественного возобновления придонецких боров. – Симферополь: ИТ «Ариал». – 2019. – 361 с.
Санников С. Н., Санникова Н. С., Кочубей А. А., Петрова И. В. Естественное возобновление сосны на гарях в лесостепи Западной Сибири // Сибирский лесной журнал. — 2019. — № 5. — С. 22-29.
Спринцын С. Н., Фрадкин А. М., Спринцын М. С. Оценка эффективности естественного возобновления сосны алеппской Pinus halepensis Mill. В Восточном Средиземноморье на примере Израиля // Сибирский лесной журнал — 2017. — № 2. — С. 3-12.
