
RARE ORCHIDS IN THE NATIONAL PARK «NIZHNYAYA KAMA» AND ADJACENT TERRITORIES IN THE NORTH OF TATARSTAN (EUROPEAN PART OF RUSSIA), THEIR HABITATS AND STATE OF POPULATIONS
JOURNAL: «EKOSISTEMY», Issue 23, 2020
Publication text (PDF):Download
UDK: 582.594(470.71):581.5
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
1.Fardeeva M. B.,
Kazan (Volga region) Federal University, Kazan, the Republic of Tatarstan, Russia
2. Lukyanova Yu. A.,
Nizhnyaya Kama National Park, Yelabuga, the Republic of Tatarstan, Russia
3.Shafigullina N. R
Kazan (Volga region) Federal University, Kazan, the Republic of Tatarstan, Russia;
Nizhnyaya Kama National Park, Yelabuga, the Republic of Tatarstan, Russia
TYPE: Article
DOI: https://doi.org/10.37279/2414-4738-2020-23-166-182
PAGES: from 166 to 182
STATUS: Published
LANGUAGE: Russian
KEYWORDS: Оrchidaceae, population, population structure, age structure, abundance, density, habitat, moss diversity.
ABSTRACT (ENGLISH):
The article examines the populations of rare species of orchids Cypripedium calceolus (L.), Epipactis helleborine L., Goodyera repens (L.) R.Br, Neottia nidus-avis (L.) Rich., Neottianthe cucullata (L.) Schlechter, Platanthera bifolia (L.) Rich. on the territory of the Republic of Tatarstan. The temperate continental climate prevails on the territory of Tatarstan, which is typical for the central part and northwestern regions of the Volga and Predkamye. In contrast, the north and northeast of Tatarstan are characterized by a moderately cool climate with a lot of precipitation during the growing season. Small fragments of the southern taiga complexes with green moss spruce and fir are formed in the north, in the Baltasi district. Dark coniferous species – spruce and fir are dominated in the north-east along the river Vyatka in Lubyanskoye forestry in coniferous-deciduous forests. Such habitats are characterized by the predominance of typically boreal and boreal-nemoral species of shrubs, grasses and mosses. Rare species have been recorded: Pyrola media Sw., P. chlorantha Sw., Diphasiastrum complanatum (L.) Holub, Hypopitys monotropa Crantz, Viola selkirkii Pursh ex Goldie. Orchid populations have some features. The development and flowering of Platanthera bifolia and Cypripedium calceolus occur 7–10 days later in comparison with central and northwestern regions of Tatarstan. Virginal and adult vegetative individuals predominate in the population structure. Generative individuals of C. calceolus in spruce-broad-leaved and pine-spruce with fir forests often do not form clones; their share is 72 % of all generative plants. The number of shoots of C. calceolus clones decreases and makes in average 3.2 shoots. On the contrary, the number of shoots in clones of the northwestern populations is equal to 6.5. The share of generative individuals of the studied populations of P. bifolia is 25 % on average and it significantly decreases in the populations of the recreational zone of the National Park. The species grows here in dry pine forests on compacted soil, and, probably, tourists pick flowers up for bouquets. Populations of N. cucullata and G. repens are found only in the protected and buffer zones of the National Park. The age structure is full-term though the share of generative plants is quite low and averages 34 % for N. cucullata, and only 15 % for G. repens. N. cucullata and G. repens have not been found yet in habitats with suitable abiotic and biotic conditions in the north and north-east of Tatarstan. This fact requires further research. The species composition of mosses is almost identical in the habitats of orchids (Pleurozium schreberi (Brid.) Mitt., dominates among mosses in all phytocenoses, but the projective cover and height of the moss tufts differ significantly. Typically boreal and hemiboreal species are registered: P. schreberi Dicranum polysetum Sw., Dicranum scoparium Hedw., Rhytidiadelphus triquetrus (Hedw.) Warnst, Rhodobryum roseum (Hedw.) Limpr., Hylocomium splendens (Hedw.) Bruch et al., Ptilium crista-castrensis (Hedw.) DeNot. A rare species – Plagiomnium drummondii (Bruch&Schimp.) T.J. Kop. is identified in the north of the Republic of Tatarstan. The abundance and development of mosses, as guarantors of humidity and preservation of microclimatic conditions, are important conditions for the growth of symbiotrophic fungi that contribute to the maintenance of orchids and many species of boreal forests on the southern border of their distribution.
Фардеева М. Б.1, Лукьянова Ю. А.2, Шафигуллина Н. Р.1, 2
1 Казанский (Приволжский) федеральный университет
Казань, Республика Татарстан, Россия
orchis@inbox.ru, nadiashafigullina@gmail.com
2 Национальный парк «Нижняя Кама»
Елабуга, Республика Татарстан, Россия
В статье рассмотрены популяционные особенности редких видов орхидей Cypripedium calceolus (L.), Epipactis helleborine L., Goodyera repens (L.) R.Br, Neottia nidus—avis (L.) Rich., Neottianthe cucullata (L.) Schlechter, Platanthera bifolia (L.) Rich. на севере Республики Татарстан. На территории Татарстана преобладает умеренно-континентальный климат, что характерно для центральной части и северо-западных районов Предволжья и Предкамья. Напротив, север и северо-восток характеризуются умеренно-прохладным климатом с большим количеством осадков в вегетационный период. На севере, в Балтасинском районе формируются фрагменты южно-таежных комплексов с елью и пихтой зеленомошных. На северо-востоке по долине реки Вятка в Лубянском лесничестве в хвойно-широколиственных лесах также преобладают темнохвойные виды – ель и пихта. Такие местообитания отличаются обилием типично бореальных и бореально-неморальных видов кустарничков, трав и мхов. Отмечены редкие виды: Pyrola media Sw., P. chlorantha Sw., Diphasiastrum complanatum (L.) Holub, Hypopitys monotropa Crantz, Viola selkirkii Pursh ex Goldie. Популяции орхидей в таких условиях имеют некоторые особенности. Развитие и цветение Platanthera bifolia и Cypripedium calceolus задерживается на 7–10 дней, по сравнению с центральными и северо-западными районами Татарстана. В популяционной структуре преобладают виргинильные и взрослые вегетативные особи. Генеративные особи C. calceolus в елово-широколиственных и сосново-еловых с пихтой лесах часто не образуют клоны, их доля составляют 72 % от всех генеративных растений. Число побегов в клонах C. calceolus сокращается и составляет в среднем 3,2 побега, напротив, в северо-западных популяциях вида число побегов в клонах в среднем составляет 6,5. Доля генеративных особей изученных популяций P. bifolia в среднем составляет 25 % и особенно снижается в популяциях рекреационной зоны Национального парка. Вид здесь произрастает в сухих сосняках на уплотненной почве, и, возможно, собирается в букеты. Популяции N. cucullata и G. repens встречаются только в заповедной и буферной зонах Национального парка. Возрастная структура полночленная, однако, численность генеративных растений довольно низкая и составляет в среднем для N. cucullata 34 %, а для G. repens всего 15 %. В подходящих по абиотическим и биотическим условиям местообитаниях на севере и северо-востоке Татарстана N. cucullata и G. repens пока не были найдены, что требует дальнейших исследований. Видовой состав мхов в местообитаниях орхидей практически идентичен (во всех фитоценозах среди мхов доминирует Pleurozium schreberi), но проективное покрытие и высота дерновинок мхов значительно различается. Отмечены типично бореальные и гемибореальные виды: P. schreberi, Dicranum polysetum Sw., Dicranum scoparium Hedw., Rhytidiadelphus triquetrus (Hedw.) Warnst, Rhodobryum roseum (Hedw.) Limpr., Hylocomium splendens (Hedw.) Bruch et al., Ptilium crista-castrensis (Hedw.) DeNot., а на севере РТ редкий вид – Plagiomnium drummondii (Bruch&Schimp.) T.J. Kop. Обилие и развитие мхов, как гарантов влажности и сохранения микроклиматических условий, являются важными условиями произрастания симбиотрофных грибов, способствующих поддержанию орхидей и многих видов бореальных лесов на южной границе их распространения.
Ключевые слова: Orchidaceae, популяция, возрастная структура, численность, плотность, местообитание, разнообразие мхов.
ВВЕДЕНИЕ
В современных условиях для снижения рисков потери биологического разнообразия и природных местообитаний редких организмов на территории Республики Татарстан последнее время проводится подготовка практических рекомендаций для лесников, основанная, в том числе, на оценке распространения редких видов растений и животных. Подобные исследования лесов и рекомендации для сохранения редких орхидей проводились в Архангельской области (Рай и др., 2017). Материал, собранный в Мортовском, Лубянском лесничествах и Национальном парке, в Балтасинском, Елабужском и Кукморском районах на севере Татарстана (РТ) рекомендовано было использовать для будущего контроля и сохранения уникальных природных комплексов при заготовках древесины Минлесхозом РТ. Важным для понимания биолого-экологических механизмов, обеспечивающих устойчивость лесных экосистем к природным и антропогенным воздействиям, является изучение популяций лесообразующих видов деревьев, собственно определяющих состав и структуру лесных фитоценозов, а также редких видов, разнообразие которых обеспечивает стабильное функционирование экосистем (Фардеева, 2014). Таким образом, исследования численности, структуры и оценки жизненности популяций редких охраняемых видов в условиях антропогенных воздействий актуальны и необходимы. В группу наиболее редких растений РТ входят виды сем. Орхидных (Orchidaceae Juss.), которых в Татарстане произрастает около 30 видов. Только дремлик широколистный, встречающийся часто по лесам республики и даже в рекреационных зонах больших городов, не внесен в Красную Книгу РТ (2016).
Причинами редкости орхидей являются специфические особенности их биологии и экологии. Растения сем. Orchidaceae имеют ряд эколого-биологических особенностей – слабое прорастание семян, микотрофность, зависимость от грибов и узкая экологическая толерантность к факторам, потому они и являются уязвимыми компонентами флоры (Вахрамеева и др., 2014). Редкость орхидных связана также с такими биологическими особенностями, как сильная редукция зародыша, отсутствие эндосперма, уменьшение размера семени, необходимость симбиоза с почвенными грибами, обеспечивающими прорастание семян, низкая конкурентная способность видов. Высокая специализация опыления является важнейшим фактором процветания и успешной репродукции орхидей, а для видов с отсутствием нектара, сложность механизмов опыления может привести к полному их исчезновению (Холодов и др., 1998; Иванов и др., 2004, 2009). Редкость орхидей обусловлена и определенными эколого-фитоценотическими требованиями – они произрастают в определенных условиях освещенности, влажности и кислотности почв, а некоторые бриофильные орхидеи обязательно приурочены к моховой подстилке. Однако, часто в нарушенных местообитаниях, где конкуренция снижена по каким-либо причинам, некоторые орхидеи могут разрастаться и захватывать большие территории.
Основной целью исследования было: оценить состояние новых местообитаний и популяций некоторых редких орхидей на севере Республики Татарстан и в Национальном парке «Нижняя Кама».
МАТЕРИАЛ И МЕТОДЫ
Согласно природному районированию РТ (Бакин и др., 2000) северные районы Татарстана, где проходили исследования – Балтасинский, Елабужский (северо-восточная часть Национального парка и Мортовское лесничество), Кукморский (Лубянское лесничество) территориально приурочены к Восточному Предкамью. Балтасинский и Кукморский районы относятся к Волжско-Вятскому возвышенно-равнинному региону темнохвойно-широколиственных неморальных лесов с фрагментами южно-таежных елово-пихтовых и сосново-еловых зеленомошных лесов, к ландшафту Правобережно-Вятского эрозионно-равнинного района подтаежных Приуральских широколиственно-пихтово-еловых и сосновых остепненных лесов. Лубянское лесничество приурочено к долинам рек Вятки и Лубянки, здесь преобладают дерново-подзолистые супесчаные почвы. Водораздельное плато рек Волги и Вятки в Балтасинском районе характеризуется обычно высотами 175–195 м. В геологическом строении рельефа участвуют породы пермской системы, преимущественно породы татарского яруса, представленные мергелями, глинами с прослоями известняков, доломитов, песчаников. Склоновые и водораздельные поверхности прикрыты суглинками.
Климат севера РТ определяется как умеренно-прохладный с достаточным увлажнением: количество осадков за период май-сентябрь составляет более 240 мм, причем в первую половину вегетационного периода (май – июнь) осадки составляют до 90 мм. Безморозный период 130 дней, снежный покров залегает в течение 150–160 дней. В целом несколько отличается от центральной части Татарстана (севера-запада и востока Предкамья РТ, в частности – Елабужского р-на и нацпарка).
Елабужский район приурочен к Вятско-Камскому равнинному региону темнохвойно-широколиственных лесов и долинных гигрофитных неморальных лесов, к ландшафту Елабужско-Предкамского эрозионно-расчлененного района подтаежных Приуральских широколиственно-пихтово-еловых и сосново-широколиственных и сосновых травяных лесов. Национальный парк «Нижняя Кама» приурочен к центральной части и востоку РТ, располагается от Восточного Предкамья до Восточного Закамья, в долине реки Камы (в Елабужском и Тукаевском районах РТ). Плотина ГЭС, перегораживающая Каму, привела к формированию огромного водного бассейна, называемого «Нижнекамским водохранилищем». Климат определяется как умеренно-континентальный, с более жарким и сухим летом и сниженным количеством осадков, в среднем до 45–55 мм осадков в вегетационный период. На данной территории климат смягчается (особенно в зимний период) за счет западного переноса воздушных масс (Переведенцев и др., 2008).
Исследования орхидей проводились в различных сообществах на севере и северо-востоке РТ: на севере были исследованы лесные фитоценозы в Балтасинском районе, на северо-востоке – Лубянском лесничестве, а также в Мортовском лесничестве и Национальном парке (НП) «Нижняя Кама». В новых местообитаниях определяли количество находок ценопопуляций орхидей, чтобы выявить не только разнообразие растительных сообществ, в которых редкие виды встречаются, но к каким фитоценозам они приурочены чаще.
В 2019 году в ходе экспедиций было проведено более 300 описаний различных фитоценозов и около 20 % содержали в видовом составе сообществ редкие виды орхидей.
Основными объектами изучения стали ценопопуляции орхидей (Orchidaceae), занесенных в Красную книгу РТ (2016): тубероидные орхидеи со сферическим клубнем: Neottianthe cucullata (L.) Schlechter – категория редкости 2 и Platanthera bifolia (L.) Rich. – категория 3; короткокорневищные виды: Cypripedium calceolus (L.) – категория 3; Epipactis helleborine L.; а также короткокорневищная орхидея с многочисленными запасающими корнями – Neottia nidus-avis (L.) Rich. – категория 3 и длиннокорневищная – Goodyera repens (L.) R.Br – категория 2.
В работе использовались общепринятые геоботанические методы и эколого-ценотический анализ местообитаний орхидей. Соотношение разных эколого-фитоценотических групп растений фитоценоза определяли в процентах от общего числа видов местообитания орхидей.
В популяционном анализе, кроме картирования ценопопуляций в фитоценозах, определялись численность, плотность, возрастная структура и базовый спектр. Возрастная структура оценивалась в процентах по каждой онтогенетической группе от общей численности популяции. Классификация состояния популяций по возрастной структуре и базовому спектру характеризовалась по Л. Б. Заугольновой (1994), как правосторонний, левосторонний или центрированный тип спектра. Также определялись коэффициент эффективности (Животовский, 2001) и индекс возрастности Δ, который определяется как (ƩKi×Δi)/ ƩKi, где Ki — численность i-той возрастной (онтогенетической) группы, Δi — «вес или индекс» возрастности i-той группы, ΣKi — численность всей популяции (Ценопопуляции…, 1976). Классификацию популяций «дельта-омега» на основе индекса возрастности и коэффициента эффективности проводили по Л. А. Животовскому (2001): молодая, переходная, зреющая, зрелая, стареющая, старая.
Базовый спектр определялся как средние значения относительной численности особей (М) каждой возрастной группы (в %), на основе изучения возрастной структуры нескольких ценопопуляций. Для базового спектра существует некоторая зона, в пределах которой возможны колебания численности – она заключается в пределах М±3σ (σ – среднее квадратичное отклонение или СКО) (Ценопопуляции…, 1976, Злобин, 2009).
Онтогенетические группы выделялись по морфологическим признакам, были определены: ювенильные (j); имматурные (im); виргинильные (v); молодые генеративные (g1); средние генеративные (g2); крайне редко – сенильные группы (s) или скорее квазисенильные растения, (проростки у орхидей не выделяются, так как ведут подземный образ жизни). Для этого использовались опубликованные онтогенезы для некоторых видов орхидей (Царевская, 1975; Вахрамеева и др., 1983; Блинова, 1993; Татаренко, 1996; Фардеева, 2002; Вахрамеева, Жирнова, 2003). Единицей популяционных исследований клубнеобразующих видов является особь, у корневищных – парциальный побег, «условная особь». Каждой особи на карте присваивались координаты (x, y), а полученные данные заносились в базу данных для дальнейшего пространственного анализа. Более подробный популяционный анализ проводился для P. bifolia и C. calceolus.
Таблица 1
Характеристики исследованных популяций на севере РТ
Примечание к таблице. Координаты центральной части участков исследования: Балтасинский р-н (д. Ципья) – N56.326990 E 050.17977; Лубянское лесничество (кв.101) – N 56.08611 E 051.46393; Мортовское лесничество – N55.94067 E 051.51479; Национальный парк (кв. 87) – N55.72107 E 052.25706.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Как отмечают большинство орхидологов (Vakhrameeva et al., 2008) стратегия размножения у орхидных разных жизненных форм отличаются: для клубневых видов (P. bifolia, N. cucullata) преимущественно отмечается семенное размножение, у корневищных (C. calceolus, E. helleborine, G. repens), как правило, преобладает вегетативное. Для клубневых орхидей характерно развитие большого числа семян с хорошо сформированным зародышем, напротив, у корневищных орхидей формируются крупные семена с мелкими зародышами. Установлено, что редкие виды орхидных являются стеновалентными и стенобионтными видами (Ишмуратова и др., 2010), то есть, приурочены к специфическим биотам. Новые местообитания орхидей, изученные на основе геоботанических описаний и видовых списков фитоценозов, были проанализированы по эколого-ценотическим характеристикам, что дало возможность разобраться, к каким преимущественно сообществам приурочены орхидеи, и выявить наиболее подходящие для них местообитания. Количество находок вида, в том или ином фитоценозе, отражено в процентах (рис. 1). По построенному спектру местообитаний, можно выявить разнообразие растительных сообществ, в которых отмечались редкие виды орхидей в Национальном парке и сопредельных территориях на севере Татарстана.
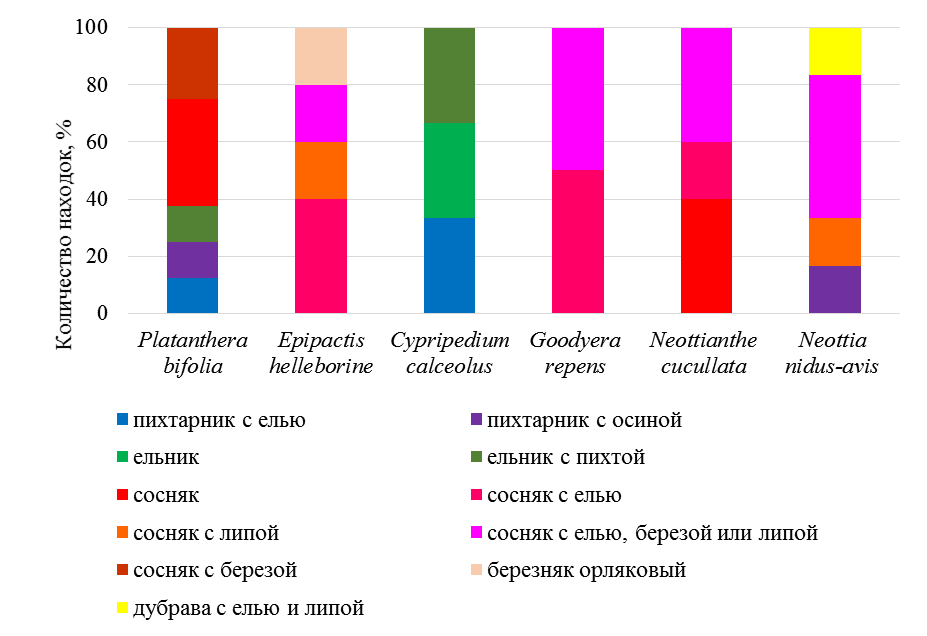
Рис. 1. Количество находок редких орхидей в различных фитоценозах на севере и северо-востоке Татарстана
Epipactis helleborine и Neottia nidus-avis
Neottia nidus—avis часто отмечается в липняках и сложных сосняках нацпарка, формируя небольшие скопления из 5–20 особей. На севере РТ – в Лубянском лесничестве и Балтасинском районе вид иногда встречается многочисленными локусами от 50 до 112 особей на 25–30 м² по мертвопокровным пихтарникам с елью лещиновым или ельникам с сосной и липой. Особенностью местообитаний гнездовки, как правило, являются участки сохранившихся старовозрастных лесов. Причем особые в них микроместообитания – наиболее затененные небольшие участки за счет разросшейся лещины или подроста липы, где под пологом сохраняется влажность и тень, почти полное отсутствие каких-либо растений, мощная подстилка разлагающейся листвы.
Из орхидей РТ самым распространенным является дремлик широколистный, который на правобережье реки Волги (Предволжье РТ) в липняках и дубравах с липой и кленом образует локалитеты до 200–400 особей. На территории нацпарка и прилегающих к нему неохраняемых участков (Шильнинский лес) он встречается в разнообразных фитоценозах – от достаточно сухих сосняков вейниковых и орляковых до сложных сосняков с липой или елью костяничных и вторичных березняков орляково-травяных. Численность ценопопуляционных локусов варьирует от 20 до 35 шт., с преобладанием до 80–90 % генеративных особей.
Neottianthe cucullata и Goodyera repens
Первоначально остановимся на характеристике местообитаний наиболее редких, так называемых бриофильных орхидеях – N.cucullata и G.repens, приуроченных к мшистым участкам. Эти таежные виды, располагая свои подземные органы по лесной подстилке и в моховом покрове хвойных и хвойно-широколиственных лесов, находятся в Татарстане на южной границе распространения, потому и очень редки. Причем если гудайера ползучая часто занимает довольно затененные и влажные участки под пологом или в тени крон елей, то неоттианте может встречаться и на более освещенных и суховатых участках сосняков орляковых и брусничных с елью или лишайниково-мшистых (Редкие виды флоры РТ, 2017). В условиях 2005–2007 годов, когда проходили специальные популяционные исследования орхидных на территории Елабужского района нацпарка, гудайера не была найдена, и долгое время не отмечалась в геоботанических описаниях лесов. Редкая орхидея, часто переходит в состояние покоя и ведет подземный образ жизни, существуя только в виде корневища за счет эксплуатации грибов, найти ее в одном и том же месте бывает сложно. Последние годы (2015, 2017, 2019 гг.) отличались высокой влажностью даже в центральной части РТ, когда среднее количество осадков в июле составляло 75–98 мм, хотя обычно составляет около 35–45 мм. Исследования 2018–2019 годов увенчались успехом и гудайера была найдена в кв. 87 и 34 в сосняках с елью зеленомошных на участке Большого Бора нацпарка.
Ценопопуляции N. cucullata и G. repens отмечались в семи местообитаниях Национального парка (рис. 2). Виды преимущественно приурочены к мшистым соснякам: сосняки лишайниково- и бруснично-мшистые с березой (кв. 87, выдел 8; 10), сосняки с елью и березой чернично- или бруснично-вейниково-мшистые (кв. 42, выдел 9; кв. 35, выдел 32), сложные сосняки с елью и липой чернично-костянично-мшистые (кв. 34, выдел 6; кв. 43, выдел 5; 6). Здесь наряду с типично бореальными и бореально-неморальными лесными видами отмечались лесо-луговые. Среди содоминантных кустарничков – Vaccinium myrtillus L., Vaccinium vitis-idaea L., Rubus saxatilis L., Orthilia secunda (L.) House и многолетних трав – Calamagrostis arundinacea (L.) Roth, Carex digitata L., Luzula pilosa (L.) Willd., Pulsatilla patens (L.) Mill. отмечались и некоторые охраняемые или редкие (внесенные в приложения к Красной книге) виды РТ. Среди них – Pyrola media Sw, P. rotundifolia L., P. chlorantha Sw., Chimaphila umbellata, (L.) Holub, Hypopitys monotropa Crantz. В сложных сосняках с липой – единично встречались Cephalanthera rubra (L.) Rich и Lilium pilosiusculum (Freyn) Miscz..
Проективное покрытие мхов в таких сообществах составляет 30–65 %, среди которых основными являются – Pleurozium schreberi (Brid.) Mitt., Dicranum polysetum Sw., Dicranum scoparium Hedw.. В меньшем количестве встречаются Rhodobryum roseum (Hedw.) Limpr., Hylocomium splendens (Hedw.) Bruch et al., которые, в условиях РТ произрастают только в лесах с значительным участием ели в древостое. Причем, G. repens развивалась непосредственно среди побегов P. schreberi, однако эти побеги не образовывали сплошного пышного мохового покрова. Схожая картина наблюдалась в другом месте произрастания G. repens на территории РТ – в Раифском участке Волжско-Камского заповедника. Возможно, большему развитию мохового покрова препятствует развитие кустарничков, таких, как черника и брусника. Неоттианте так же росла вблизи крупных моховых куртин, в основном среди разреженных побегов P. schreberi.
В популяционной структуре G. repens преобладали вегетативные растения, представленные двух- и 4–5-листными розетками, образуя несколько компактных куртин, среди которых было обнаружено 5 генеративных побегов. Численность разных возрастных групп составляет – 4:23:5 (im: v: g), преобладают виргинильные розетки.
Численность ценопопуляций (ЦП) N. cucullata на учетных площадках, размером 50 м² варьировала от 56 особей в сосняках бруснично-мшистых с березой (кв. 87) и 79 особей в сосняках с елью и березой бруснично-вейниково-мшистых (кв. 43), до 121 особей в сосняках с елью и липой чернично-костянично-мшистых (кв. 34). Возрастные спектры ценопопуляций аналогичны, имеют полночленную структуру, доля виргинильных и молодых генеративных особей почти одинакова, доля ювенильных и имматурных – довольно высокая. Общий тип базового спектра в процентах составляет 18: 21: 27: 26: 8 (j: im: v: g1: g2).
Хорошее проективное покрытие мхов в исследованных местообитаниях поддерживает влажность даже в сухих светлых сосняках и сосняках с березой на территории Большого Бора Национального парка. Эта особенность местообитаний – сохранность и развитие мохового покрова, способствует развитию симбиотрофоных грибов, так необходимых для многих редких бореальных видов – грушанок, подъельника, орхидей.

Рис. 2. Neottianthe cucullata (a) и Goodyera repens (b) в Национальном парке«Нижняя Кама» (Фото Ю. Лукьяновой)
Platanthera bifolia и Cypripedium calceolus
Platanthera bifolia и Cypripedium calceolus по территории РТ встречаются довольно часто, находясь здесь в центре ареала. Однако, высокодекоративные растения часто собираются в букеты или выкапываются для посадки.
Platanthera bifolia
Лесо-луговая любка двулистная встречается по хвойным, хвойно-широколиственным лесам, опушкам, полянам, вдоль просек, окраинам болот и реже по светлым окнам в широколиственных сообществах правобережья реки Волги.
Вид обладает наибольшим диапазоном экологической валентности по отношению к факторам освещенности и влажности биотопов, по сравнению с другими видами, что обуславливает широкий характер ее распространения, как в Национальном парке, так и на сопредельных неохраняемых участках на севере РТ в Мортовском, Лубянском и Балтасинском лесничествах (рис. 3). В основном любка двулистная как на северо-западе в притерассных понижениях реки Волги, так и в нацпарке по долине реки Камы встречается в светлых сосняках, березняках, сосняках с березой вейниково-орляковых, бруснично- или чернично-мшистых и разнотравно-костяничных, что составляет 62,5 % от всех выявленных местонахождений, где преобладает боровое и бореально-неморальное разнотравье, а иногда лесо-луговые и лесо-степные виды (рис. 4).
 |
 |
Рис. 3. Platanthera bifolia в Национальном парке «Нижняя Кама» (Фото Ю. Лукьяновой)
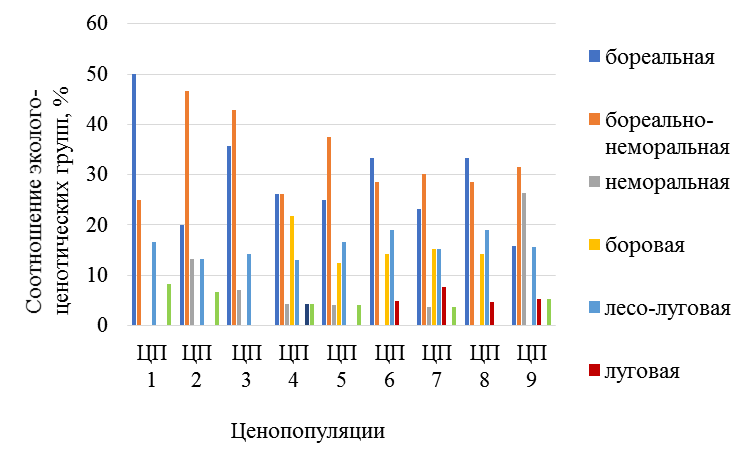
Рис. 4. Соотношение эколого-ценотический групп растений в местообитаниях ЦП Platanthera bifolia на севере и северо-востоке РТ
На севере РТ (Балтасинский р-н) P. bifolia отмечалась в пихтарниках с осиной мшистых или пихтарниках с липой травяных, а также ельниках с пихтой зеленомошных, на более затененных участках (37,5 % местонахождений), часто, располагая свои листья на моховых дерновинках. В подобных лесах преобладает бореальное и бореально-неморальное разнотравье, реже неморальное, что характерно для хвойно-широколиственных лесов севера РТ. Однако P. bifolia в елово-пихтовых и елово-широколиственных лесах была приурочена к особым микроместообитаниям – это участки поваленных деревьев и прогалы, где полог хвойного леса более разреженный.
Приоритетным направлением Национального парка является выявление численности и состояния популяций редких видов, в связи с чем, проводится постоянный мониторинг известных местонахождений и поиск новых. Как правило, в Национальном парке редкие виды орхидей (P. bifolia, N.cucullata, G.repens) встречаются преимущественно в заповедной и буферной зонах, где снижена рекреация и сохраняются зональные бореальные комплексы.
В 2005–2006 годах любка двулистная в нацпарке часто встречалась по влажным и светлым березнякам с елью сфагновым, которые представляют собой вторичные сообщества, возникшие на месте рубок и пожаров ельников (кв. 25, 26). Современные исследования (2018–2019 гг.) в основном проводились на территории «Большого Бора», в сосняках. В исследованных местах произрастания P. bifolia отмечается обильное развитие мохового покрова, который часто представлен сплошными куртинами, с проективным покрытием 30–50 %. Такая картина наблюдается в условиях хорошего освещения, которое бывает на опушках, прогалах в местах выпавшего древостоя, в сосновых посадках нацпарка. Доминируют P. schreberi, D. polysetum, D. scoparium. На севре РТ в Балтасинском р-не проективное покрытие мхов в пихтарниках с елью увеличивается до 90–100 %, в основном это P. schreberi, D. scoparium, а также Rhytidiadelphus triquetrus (Hedw.) Warnst., Rh. roseum, H. splendens.
Для анализа численности и возрастной структуры P. bifolia были проанализированы четыре ценопопуляции в Национальном парке (ЦП 1–4), и на неохраняемой территории – Лубянского (ЦП 5, ЦП 6), Мортовского (ЦП 7) лесничеств и Балтасинского р-на (ЦП 8 и ЦП 9). Все популяции произрастают в местообитаниях, которые отличаются по типу фитоценоза и степени нарушенности.
В Большом Бору нацпарка плотность особей в популяциях совместима с плотностью на севере РТ и в среднем для разных возрастных групп составляет 0,4–0,6 ос. на 1м², общая плотность варьирует от 1,8 до 2,3. Напротив, в рекреационной зоне нацпарка и Шильнинском лесу (ЦП3 и ЦП4) плотность разных возрастных групп варьирует от 0,08 до 0,2. Причиной часто является низкое проективное покрытие трав и снижение мохового и лишайникового покрова, а также уплотненность и задерненность почвы луговым разнотравьем, либо в сосняках с липой – разрастание широкотравья (рис. 3).
Популяции P. bifolia в Лубянском лесничестве небольшие, любка не образует здесь крупных скоплений: численность ЦП 5 составляет 43 особей, численность ЦП 6 – 29 особей, что обусловлено затенением и разрастанием кустарничков в сосняках с елью чернично-костянично-мшистых. Напротив, в Мортовском лесничестве в светлом сосняке бруснично-мшистом и Балтасинском р-не в прогале пихтарника зеленомошного встречаются крупные популяции вида, где на площади 60–70 м² отмечается 135 (ЦП7) и 110 (ЦП8) особей. В затененных ельниках с липой лещиновых (ЦП9) численность снижается до 21 особей на 20 м².
Возрастные и базовые спектры ценопопуляций, как в нацпарке, так и на сопредельных неохраняемых участках – полночленные, левосторонние, с преобладанием прегенеративных групп (рис. 5–8). В светлых сосняках нацпарка – Большой Бор, Танаевский и Шильнинский лес, Мортовское лесничество возобновление P. bifolia, лучше – доля ювенильных и имматурных растений составляет 45–70 %. Напротив, в условиях затенения сосново-еловых и, особенно елово-пихтово-широколиственных лесов (Лубянское и Балтасинское лес-ва) их доля сокращается до 25–45 %, иногда ювенильные растения отсутствуют вовсе (ЦП9).
На территории нацпарка численность и плотность генеративных растений P. bifolia варьирует от 26 до 42 %, однако резко снижается в рекреационной зоне (ЦП3) до 12 %, причем зрелые генеративные группы вообще не отмечаются, что обусловлено, возможно, сбором цветов в букеты (рис. 5–6).
На севере и северо-востоке Татарстана (Мортовское, Лубянское, Балтасинское лесничества) в сосновых, сосново-еловых и елово-пихтовых лесах доля генеративных особей
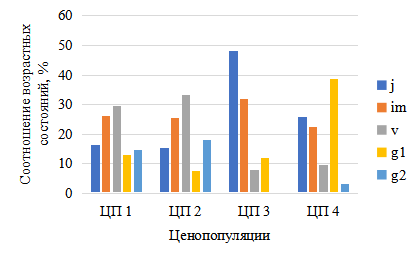
Рис. 5. Возрастная структура ценопопуляций Platanthera bifolia в Национальном парке
Возрастные группы: ювенильные (j); имматурные (im); виргинильные (v); молодые генеративные (g1); зрелые генеративные (g2).
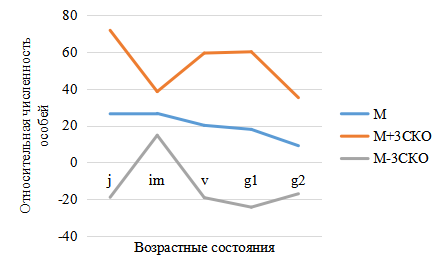
Рис. 6. Базовый спектр ценопопуляций Platanthera bifolia в Национальном парке
Обозначения: М – средние значения относительной численности особей каждой возрастной группы (в %); М±3СКО (среднее квадратичное отклонение).
в возрастной структуре популяций P. bifolia сокращается в среднем до 25–30 %. Увеличивается доля виргинильных растений, в которую часто входят взрослые вегетативные особи, представляющие собой временно нецветущие генеративные растения. Что хорошо отражено в базовом спектре (рис. 8), представляющем собой средние значения относительной численности особей 5-ти исследованных ценопопуляций. На основе определения возрастных групп были рассчитаны индекс восстановления, коэффициенты возрастности и эффективности, отражающие процессы восстановления, возраст популяций и эффективность использования природных ресурсов (рис. 9).
В целом можно отметить, что индексы восстановления везде довольно высокие и варьируют от 0,55–0,88, то есть за счет интенсивного семенного возобновления в ЦП P. bifolia на 55–88 % восстанавливается генеративная фракция. Анализируя коэффициенты возрастности и эффективности, по предложенной Л. А. Животовским (2001) классификации «дельта-омега», можно отметить, что все ценопопуляции P. bifolia – молодые (рис. 9).
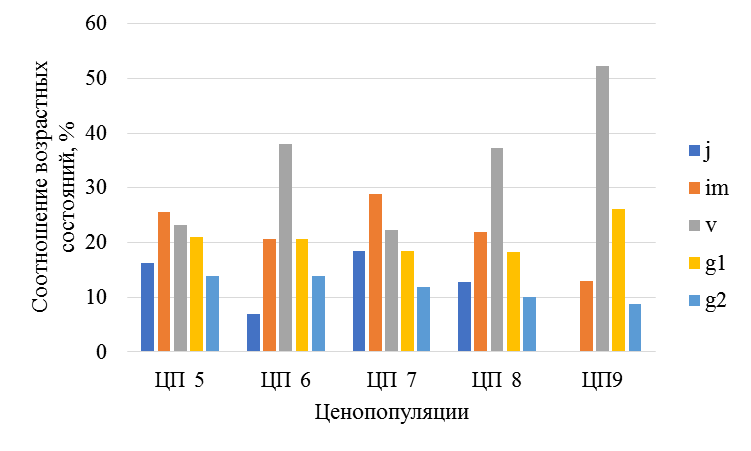
Рис. 7. Возрастная структура ценопопуляций Platanthera bifolia на севере Татарстана
Возрастные группы: ювенильные (j); имматурные (im); виргинильные (v); молодые генеративные (g1); зрелые генеративные (g2).
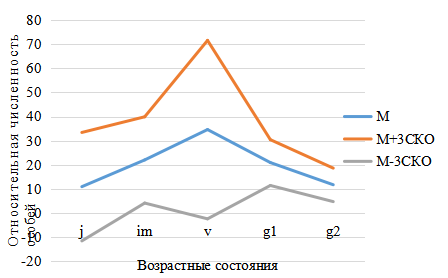
Рис. 8. Базовый спектр ценопопуляций Platanthera bifolia
Cypripedium calceolus
Впервые для РТ в 2017–2019 годы C. calceolus был найден на трех участках в елово-широколиственных неморальных и елово-сосново-пихтовых разнотравно-мшистых лесах, на водораздельном плато между реками Волгой и Вяткой, где в строении рельефа участвуют породы пермской системы – мергеля, глины с прослоями известняков и доломитов, почвы суглинистые, богатые минеральными веществами. Балтасинский район РТ граничит с Кировской областью и республикой Марий Эл. На сопредельных с Татарстаном территориях в Кировской области изучение ЦП C. calceolus проводилось в окрестностях населенного пункта Пашино. Вид встречается «в еловых, елово-пихтовых и сосново-еловых лесах на известняках, а также в заболоченных осиново-еловых лесах на торфянистых почвах» (Харина, 2019). В Марий Эл ценопопуляции C. calceolus отмечаются по березнякам с елью заболоченным и березнякам лишайниковым (Попугаева и др., 2016).
В Балтасинском районе в 3-х местообитаниях C. calceolus преобладают бореальные виды – Oxalis acetosella L., Gymnocarpium dryopteris (L.) Newman, Viola selkirkii Pursh ex Goldie,
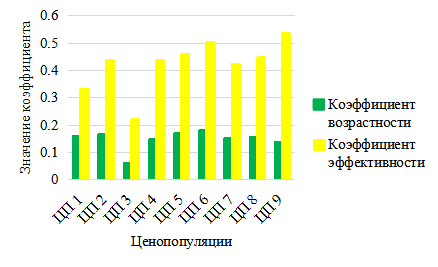
Рис. 9. Коэффициенты возрастности и эффективности ЦП Platanthera bifolia
Maianthemum bifolium (L.) F.W. Schmidt, P. rotundifolia, P. chlorantha; бореально-неморальные и неморальные виды растений – Crepis sibirica L., Aconitum excelsum Rchb., C. digitata и C. rhizina Blytt ex Lindblom, Pulmonaria obscurea Dumort., Viola mirabilis L., Milium effusum L., Aegopodium podagraria L., Bupleurum aureum Fisch. ex Hoffm. (рис.10).
Моховой покров хорошо развит, местами обилен, представлен бореальными и гемибореальными видами: P. schreberi, D. scoparium, D. polysetum, Rhytidiadelphus triquetrus (Hedw.) Warnst., Rh. roseum, H. splendens, Ptiliumcrista-castrensis (Hedw.) DeNot., Plagiomnium cuspidatum (Hedw.) T. Kop. В отличие от других фитоценозов, здесь наблюдался редкий для РТ вид – Plagiomnium drummondii (Bruch&Schimp.) T.J. Kop. Он включен в Красную книгу РТ с категорией редкости – 2 (вид, сокращающий численность). Плагиомниум Драммонда распространен на территории средней части европейской России в подзонах южной и средней тайги, где встречается единично, на гнилой древесине, почве, в основании стволов, во многих областях известен по единичным находкам (большей частью старым) (Флора мхов России, 2018). Климатические условия на севере РТ, с более низкими летними температурами и большим количеством осадков способствуют видовому разнообразию и более быстрому росту мхов (Шафигуллина и др., 2019).
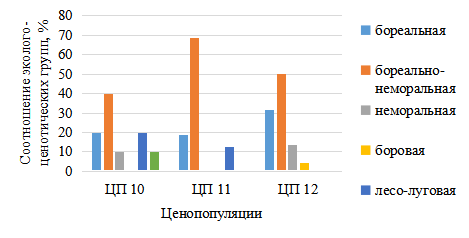
Рис. 10. Соотношение эколого-ценотический групп растений в местообитаниях ЦП Cypripedium calceolus на севере Татарстана
Популяции C. calceolus приурочены в Балтасинском районе к различным фитоценозам ЦП10: (зарастающая вырубка в пихтарнике с елью и сосной зеленомошном), ЦП11: (склон оврага в ельнике с липой кислично-мшисто-травяном), ЦП12: (ельник с липой кислично-снытевый). Численность трех ЦП варьирует от 16 до 95 особей – наибольшая отмечена на освещенной зарастающей вырубке в пихтарнике с елью и сосной зеленомошном (ЦП 10), плотность разных возрастных групп варьирует от 0,86 до 2,3 особей на 1 м², причем наибольшая плотность характерна для виргинильных и взрослых вегетативных растений и составляет 0,4–0,35. Башмачок настоящий часто разрастается на определенной стадии лесовосстановительной сукцессии сообществ, что обусловлено хорошей освещенностью и сниженной межвидовой конкуренцией с лесным разнотравьем, именно такие условия и сформировались на зарастающей вырубке. А наличие обильного мохового покрова на севере способствует сохранению влаги. Напротив, в ельниках с липой травяных (ЦП11 и 12) численность его снижена из-за сильного затенения, и интенсивного разрастания сныти и осоки волосистой, здесь он сохраняется по склонам овражков.
Анализ возрастных спектров ценопопуляций C. calceolus показал, что, в течение ряда лет (2017–2019 гг.) популяции имеют полночленную структуру, правосторонний тип с преобладанием вегетативных и генеративных особей.
В целом подобные возрастные спектры характерны для корневищных видов со сложным онтогенезом и неполным омоложением, что обусловлено вегетативным размножением, при этом процент ювенильных и имматурных особей небольшой (рис. 11).
Правосторонний тип возрастной структуры и базового спектра отражает устойчивость ценопопуляций. Базовый спектр популяции C. calceolus Балтасинского района РТ определялся как средний за 3 года в разных ценопопуляциях, он имеет центрированный тип (рис.12). Генеративных растений мало, процессы цветения на севере снижены. Максимум в этом спектре приходится на виргинильные и взрослые вегетативные особи, минимум в спектрах характерен для ювенильных и имматурных групп, что обусловлено редкостью семенного размножения C. calceolus, так как успехи репродукции и прорастание семян зависят от опылителей и микоризообразующих грибов, активность которых, возможно на севере РТ снижена.
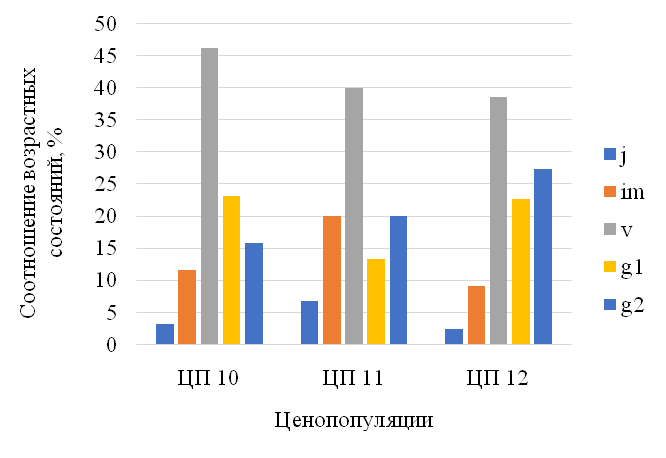
Рис. 11. Возрастная структура ценопопуляций Cypripedium calceolus
Возрастные группы: ювенильные (j); имматурные (im); виргинильные (v); молодые генеративные (g1); зрелые генеративные (g2).
Отмечено, что в популяциях C. calceolus на севере РТ, число куртин снижается – всего 7–8 куртин на 100 м², по сравнению с северо-западными ценопопуляциями в ООПТ «Семиозерка», где на 100 м² в среднем отмечается 25–30 куртин. Число побегов в куртинах
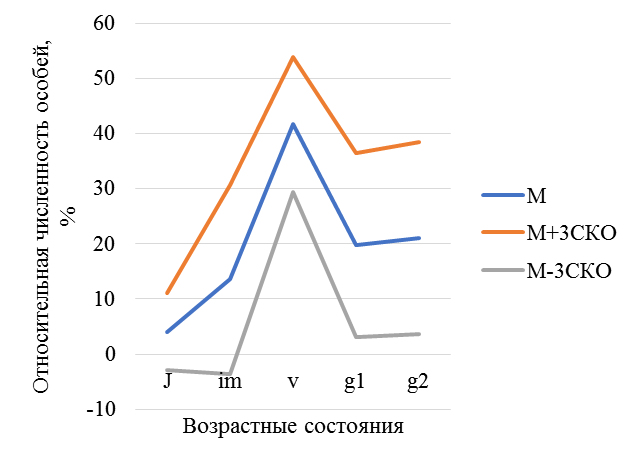
Рис. 12. Базовый спектр ценопопуляций Cypripedium calceolus
М – средние значения относительной численности особей каждой возрастной группы (в %); М±3СКО (среднее квадратичное отклонение).
(клонах) небольшое 2–3 шт., максимально 5, среднее число побегов в куртине на севере РТ – 3,2, на северо-западе – 6,5, максимально 12 побегов. На севере РТ часто отмечаются одиночные генеративные особи, не приступившие к формированию куртин, которые составляют 72 % от всех генеративных побегов.
В популяциях башмачка настоящего на территории Кировской области (Харина, 2019) и Марий Эл (Попугаева и др., 2016) также преобладают виргинильные и генеративные особи, которые образуют немногочисленные куртины. Подобную закономерность только на южной границе ареала C. calceolus отмечает В. В. Фатерыга (2019) в Крыму, где вид встречается на лесной прогалине с участием Fagus sylvatica L. и Pinus sylvestris L.
На основе показателей возрастности и эффективности по классификации Л. А. Животовского (2001) две ценопопуляции (ЦП10–11) являются «нормальными», в состоянии переходного типа. Напротив, в условиях сильного затенения ЦП C. calceolus (ЦП12), произрастающая в ельнике с липой – молодая, так как генеративных особей крайне мало (рис. 13). Цветение C. calceolus на северо-западе Татарстана начинается 24–26 мая и продолжается до 8–10 июня. На севре РТ цветение задерживается на неделю и начинается 3–6 июня и продолжается до 18–20 июня, причем в затененных участках – под пологом елей и пихт, например, 6.06.2018 и 10.06.2020 еще отмечались генеративные особи башмачка настоящего только в стадии развертывания бутонов (рис. 14).
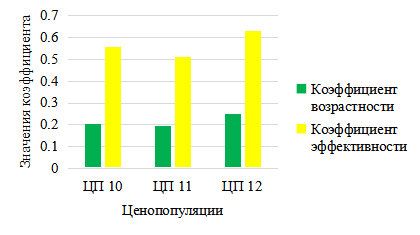
Рис. 13. Коэффициенты возрастности и эффективности ЦП Cypripedium calceolus

Рис.14. Cypripedium calceolus в Балтасинском р-не Республики Татарстан (Фото Н. Шафигуллиной)
ЗАКЛЮЧЕНИЕ
Несмотря на то, что видовой состав мхов в местообитаниях орхидей практически идентичен (во всех фитоценозах среди мхов доминирует P. schreberi) проективное покрытие и высота дерновинок существенно различаются для разных видов орхидей. Различия в обилии и развитии мхов являются важными условиями для распространения орхидей. С одной стороны, большинству исследованных видов орхидей необходимо присутствие мхов как гаранта влажности и сохранения микроклиматических условий для развития микоризообразующих грибов, способствующих также поддержанию жизнедеятельности большинства редких видов вересковых.
С другой стороны, моховой покров, по-видимому, необходим и для прорастания семян лесообразующих видов – ели и пихты, что в целом способствует сохранению типичных бореальных лесов в пограничных условиях. К сожалению, в подходящих местообитаниях для произрастания бриофильных видов G. repens и N. cucullata в Балтасинском и Лубянском лесничествах, данные виды найдены не были, что, возможно, связано с периодическими рубками лесов и требует дальнейших исследований этой территории.
В целом популяционная структура P. bifolia и C. calceolus в северных районах РТ имеют некоторые отличия от структуры их популяций в центральной части (нацпарк) и на северо-западе РТ. В условиях умеренно-прохладного климата с большим количеством осадков вегетационного периода на севере РТ, в елово-широколиственных и сосново-пихтово-еловых зеленомошных лесах развитие видов задерживается на 7–10 дней. В возрастной структуре P. bifolia и C. calceolus преимущественно отмечаются вегетативные особи. В ЦП C. calceolus заметно снижение куртинообразования в онтогенезе генеративных особей и в основном развитие одиночных генеративных растений. Изменение показателей возрастности и эффективности за 1–2 года исследований не столь информативны, только в динамике за 10–15 лет, на основе данных характеристик и показателя «дельта-омега» можно проследить цикличность популяционной динамики.
Благодарности. Авторы выражают признательность к.б.н., доц., Шайхутдиновой Г. А. (Казанский федеральный университет), а также работникам лесничеств Минлесхоза РТ в проведение экспедиций на севере Татарстана. Лично к.б.н. Аськееву О. В. (Институт проблем экологии и недропользования АН РТ, г. Казань) за помощь в поиске орхидей у села Ципья Балтасинского р-на РТ.
Работа частично выполнена в рамках НИР: Подготовка определителя ключевых биотопов и объектов, подлежащих сохранению при освоении лесосек на территории Республики Татарстан, для нужд Министерства лесного хозяйства Республики Татарстан.
REFERENCES
Бакин О. В., Рогова Т. В., Ситников А. П. Сосудистые растения Татарстана. – Издательство Казанского университета, 2000. – 496 с.
Блинова И. В. Эколого-биологические особенности Platanthera bifolia (L.) Rich. на северной границе ареала // Флористические и геоботанические исследования в Мурманской области (сб. научн. ст.) / [Ред. Н. А. Константинова]. – Апатиты, 1993. – С. 107–113.
Вахрамеева М. Г., Варлыгина Т. И., Татаренко И. В. Орхидные России (биология, экология и охрана): монография. – М.: Товарищество научных изданий КМК, 2014. – 437 с.
Вахрамеева М. Г., Денисова Л. В. Любка двулистная (Plantanthera bifolia (L.) Rich.) // Диагнозы и ключи возрастных состояний луговых растений / [Ред. Г. И. Серебряков]. – М.: Издательство Московского государственного педагогического института им: В. И. Ленина, 1983. – C. 16–18 с.
Вахрамеева М. Г., Жирнова Т. В. Неоттеанте клобучковая // Биологическая флора Московской области. – 2003. – Вып. 15. – С. 50–61.
Животовский Л. А. Онтогенетические спектры, эффективная плотность и классификация популяций // Экология. – 2001. – № 1. – С. 3–7.
Заугольнова Л. Б. Структура популяций семенных растений и проблемы их мониторинга: автореферат дисс. … д-ра биол. наук в форме научного доклада. – СПб: Санкт-Петербургский государственный университет, 1994. – 70 с.
Злобин Ю.А. Популяционная экология растений: современное состояние, точки роста: монография. – Сумы: Университетская книга, 2009. – 263 с.
Иванов С. П., Холодов В. В. Анализ характера опыления безнектарных орхидей (Orchidaceae) в зависимости от их пространственного размещения // Вопросы развития Крыма (Научн.-практич. дискус.-аналит. сб.). – Симферополь: Таврия-Плюс, 2004. – Вып. 15: Проблемы инвентаризации Крымской биоты. – С. 57–65.
Иванов С. П., Фатерыга А. В., Тягнирядно В. В. Эффективность опыления орхидей (Orchidaceae), цветущих одиночно и группами // Бюллетень Никитского ботанического сада. – 2009. – Вып. 98. – С. 22–26.
Ишмуратова М. М., Набиуллин М. И., Суюндуков И. В., Ишбирдин А. Р. Орхидеи Башкирского заповедника и сопредельных территорий. – Уфа: Гилем, 2010. – 175 с.
Красная Книга Республики Татарстан (животные, растения, грибы) / [Гл. ред. А. А. Назиров]. – Казань: Идел-Пресс, 2016. – 760 с.
Переведенцев Ю. П., Шерстюков Б. Г., Наумов Э. П., Верещагин М. А., Хабутдинов Ю. Г., Исмагилов Н. В., Тудрий В. Д. Основные особенности климата последних десятилетий на территории Татарстана // Ученые записки Казанского государственного университета. Естественные науки. – 2008. – Т. 150, кн. 4. – С. 21–32.
Попугаева Т. А., Османова Г. О. Онтогенетическая структура ценопопуляций Башмачка настоящего Cypripedium calceolus L. в окрестностях деревни Коркатово // Альманах мировой науки. – 2016. – № 2-1 (5). – С. 23–24.
Рай Е. А., Бурова Н. В., Амосова И. Б., Паринова Т. А., Пучнина Л. В., Сидорова О. В. Руководство по сохранению орхидей при заготовке древесины в Архангельской области – Архангельск: Всемирный фонд дикой природы(WWF), 2017. – 164 с.
Редкие виды флоры Республики Татарстан / [Пред. ред. сов. Ф. С. Абдулганиев] – Казань: Издатель Сергей Бузукин, 2017. – 271 с.
Татаренко И. В. Орхидные России: жизненные формы, биология, вопросы охраны. – М.: Аргус, 1996. – 207 с.
Фардеева М. Б. Онтогенез Cypripedium calceolus L. // Онтогенетический атлас лекарственных растений. – Йошкар-Ола, 2002. – С. 114–120.
Фардеева М. Б. Экологические и биоморфологические закономерности пространственно-онтогенетической структуры популяций растений, динамика и мониторинг: автореф. дис. … д-ра биол. наук. – Казань: Казанский федеральный университет, 2014. – 45 с.
Фатерыга В. В. Современное состояние ценопопуляции Cypripedium calceolus (Orchidaceae) в Крыму // Экосистемы. – 2019. – Вып. 20. – С. 76–81.
Флора мхов России. Том 4. Bartramiales – Aulacomniales / [Отв. ред. М. С. Игнатов] – М: Товарищество научных изданий «КМК», 2018. – 543 с.
Харина В. А., Пересторонина О. Н. Оценка состояния ценопопуляции Cypripedium calceolus L. на северо-востоке Кировской области // Биодиагностика состояния природных и природно-техногенных систем: Матер. ХVII Всероссийской научно-практической конференции с межд. участием (г. Киров, 2-5 декабря 2019 г.). – Киров, 2019. – С. 203–207.
Холодов В. В., Назаров В. В., Иванов С. П. Видовой состав пчел (Hymenoptera, Apoidea) опылителей некоторых видов орхидей в Крыму // Экосистемы Крыма их оптимизация и охрана (Тематич. сб. научн. тр.). – Симферополь: СГУ, 1998. – Вып. 10. – С. 81–86.
Царевская Н. Г. Любка двулистная // Биологическая флора Московской области. – 1975. – Вып. 2. – С. 11–17.
Ценопопуляции растений: основные понятия и структура / [Отв. ред. А. А. Уранов]. – М.: Наука, 1976. – 217 с.
Шафигуллина Н. Р., Каржавкина Е. Н., Зиятдинова З. Ф. Влияние различных факторов на продуктивность мохового покрова хвойных лесов Республики Татарстан // Экосистемы. – 2019. – Вып. 17 (47). – С. 44–55.
Vakhrameeva M. G., Tatarenko I. V., Varlygina T. I. et al. Orchids of Russia and adjacent countries (within the borders of former USSR) – Liechtenstein: A.R.G. Gantner Verlag Ruggell, 2008. – 690 p.
