
THE INFLUENCE OF THE BURROWING ACTIVITY OF THE WILD BOAR (SUS SCROFA L.) ON HUMUS SUBSTANCES IN SOILS
JOURNAL: «EKOSISTEMY», Issue 21, 2020
Publication text (PDF):Download
UDK: 57.025:599.731.1:630*114.34
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
- Pilipko E. N., Vologda State Dairy Farming Academy named after N.V. Vereshchagin, Vologda, Russia
- Pakhomov A. E., Oles Honchar Dnipro National University, Dnipro, Ukraine
- Toropova P. V., Vologda State Dairy Farming Academy named after N.V. Vereshchagin, Vologda, Russia
TYPE: Article
DOI: https://doi.org/10.37279/2414-4738-2020-21-142-148
PAGES: from 123 to 141
STATUS: Published
LANGUAGE: Russian
KEYWORDS: wild boar, habitat-modifying activity, burrowing activity, humus substances, carbon of humic and fulvic acids.
ABSTRACT (ENGLISH):
The burrowing activity of the wild boar (Sus scrofa L.) as one of the types of the habitat-modifying activity of terrestrial mammals has been considered. The typical species and the area of the burrowing sites in winter and summer, as well as the dynamics of humus substances are given. The literature indicates that loosening and mixing the upper soil layers cause changes in the chemical characteristics of the soil, disturb vegetation, and reduce evaporation from the soil surface, which leads to a change in the hydrothermal regime of edaphotop and an increase in both the soil climate continentality and the active layer of edaphotop. Soils of different granulometric composition were considered for comparative characteristic: loams and loamy sands in different geographical zones – steppe (the Dnipropetrovsk region, Ukraine) and taiga (wood sorrel spruce forest, Gryazovets district of the Vologda region, Russia). The steppe zone is dominated by blacktop soils, while podzolic soils prevail in the taiga zone. Although the primary characteristics of the soils were different, the tendency of changing the humus substances content in the soils under the wild boars’ burrowing sites had similar qualities. The nature of the humus substances dynamics depends on the decomposition period of organic substances left on the burrowing sites in the form of wild boar’s excreta and plant food residues. Further mineralization of dead plant remains of damaged roots of tree and shrub vegetation resulting from loosening and trophic activity of wild boar occurs on fellings after spruce indigenous forests. Part of the plant remains on the surface of the loosened soil, and the other part in the soil. As a result of the conducted researches the positive functional impact of the wild boar on any types of biogeocenoses due to its burrowing activity was confirmed.
ВВЕДЕНИЕ
Одним из наиболее важных условий существования биогеоценоза как экологической системы является эффективное взаимодействие всех составляющих её компонентов. К таким компонентам относится зооценоз, который посредством средообразующей (средопреобразующей) деятельности оказывает серьёзное влияние на устойчивость и жизнеспособность экосистемы. Механизм воздействия наземных млекопитающих на среду обитания – один из самых интересные и актуальных вопросов экологии.
Роющая деятельность млекопитающих относится к механическому типу воздействия, роющему классу деятельности и копательно-разрыхляющему виду средопреобразующей деятельности по классификации В. Л. Булахова (Булахов, 1973). Механический тип воздействия связан с нарушением почвенного и напочвенного покрова и вызывает резкое изменение физико-химических условий почвы и особый терриогенный аэрогидротермический и химический режим почвы (Абатуров, 1972, 1984; Злотин, Ходашова, 1973; Злотин, 1975; Пахомов, 1998а, 1998б; Пилипко, 2015, Пилипко Е., Пилипко А., 2015).
Дикий кабан (Sus scrofa L.) относится к типичным животным – почвороям, которые нарушают поверхностый почвенный покров вследствие трофической необходимости, то есть в результате поиска пищи. Перерывая и разрыхляя почвенный покров, кабан способствует изменению твердости почвы, что в значительной степени сказывается на ее теплопроводности. Кроме того, в результате механического воздействия на поверхность почвы, нарушается растительный покров, что способствует нагреву почвы и уменьшению испарения влаги с ее поверхности. Все это, в результате, приводит к изменению гидротермического режима эдафотопа, которое основывается не только на разрушении напочвенного покрова, а и естественного сложения почв под териогенным воздействием. Это способствует повышению меры континентальности почвенного климата и увеличению деятельного слоя эдафотопа, что, несомненно, оказывает положительное воздействие на генезис исходных почвогрунтов (Пахомов и др., 2003).
За одну кормежку кабан поедает 2–3 кг различного корма. Ни одно наземное животное не оказывает такого масштабного влияния на почву и растительный покров лесов и лугов, как дикие свиньи, переворачивающие огромную массу поверхностных слоев почвы. В отношении почвенной (насекомые, подземная фитомасса) и напочвенной (надземная фитомасса) трофики у кабана практически нет конкурентов среди млекопитающих, за исключением крота. Также важно учитывать, что в количественном отношении выявлено значительное преобладание пищевых ресурсов в почвах перед надземными кормами.
Цель исследований – динамика необходимых для поддержания плодородия почв гумусовых веществ под влиянием роющей деятельности кабана как естественного механизма, способствующего гомеостазу эдафотопа и устойчивости экосистемы.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводились в разных географических зонах – в биогеоценозах Степного Приднепровья (Центральная часть Украины) и в таёжных биогеоценозах Вологодской области (Северо-Запад России). Исследования проводились в период 2014–2018 годов. При проведении исследований в качестве методического руководства были использованы работы многих учёных (Формозова, 1950, 1952; Новикова, 1953; Данилов и др., 1966; Матюшкина, 1977; Челинцева, 1985). На учетных маршрутах во все сезоны года фиксировались следы жизнедеятельности кабанов: отпечатки следов копыт животных на почве в местах единичной встречаемости следов и натоптанных миграционных троп, экскременты, порои и непосредственные визуальные встречи с самим животным. Порои кабана изучались с помощью методик, предложенных А. Е. Пахомовым (1998, 1998а).
С целью выяснения особенности динамики важных гумусовых веществ, влияющих на плодородие почвы под влиянием роющей деятельности кабана дикого, были отобраны почвенные пробы повреждённого слоя глубиной до 20 см (ГОСТ 28168-89). Химический анализ почвы, отобранной вблизи биостационара в Присамарье Днепропетровской области, выполнялся в аккредитованной лаборатории «Химия почв» в научно-исследовательском институте биологии при Днепропетровском национальном университете им. Олеся Гончара. Анализ почв таёжных биогеоценозов Вологодского региона проводился в аккредитованной испытательной лаборатории Федерального государственного учреждения «Государственный центр агрохимической службы «Вологодский». В обеих лабораториях применялись стандартные методики: общее содержание органического углерода определялось по методикам И. В. Тюрина (1965), групповой состав гумуса почв – по методике М. М. Кононовой и Н. П. Бельчиковой (1950).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Для сравнительной характеристики было рассмотрено влияние роющей деятельности кабана дикого на гумусовые вещества в почвах двух разных географических зон – в степной, где преобладают чернозёмы и в таёжной, для которых характерны почвы подзолистого ряда.
Как в Вологодской, так и в Днепропетровской областях встречаются практически все виды повреждения почвы и напочвенного покрова в результате роющей деятельности кабана дикого из классификации Данилкина (2002) – поверхностные, сплошные, точечные и диффузные.
Порои кабана имеют сезонные отличия. Зимние, как правило, точечные и диффузные. В начале зимы наблюдается промерзание почвы, а во второй половине зимы высокая мощность снежного покрова, поэтому порои не могут быть масштабными. Масштабность таких пороев занимает всего 2–4 % на 100 м2.
В летний период встречается самое большое разнообразие почвенных пороев как по форме, так и по масштабам и глубине.
Самые распространённые формы – диффузные, на их долю приходится около 72 % от всех зафиксированных пороев, далее точечные – 11 %, поверхностые – 4 % и сплошные – 13 %. Глубина пороев в летний период от 0 до 34 см. Масштаб сплошных пороев на отдельных территориях может достигать 60 % на 100 м2.
В первую половину весеннего периода порои чаще всего диффузые и поверхностые, так как почва ещё не достаточно прогрелась и не оттаяла, но во второй половине весны порои приобретают глубокий и масштабный характер. На высокую интенсивность пороев в конце весны в Днепровском Присамарье также указывал А. Е. Пахомов (1998). На осень приходится самая высокая сезонная интенсивность пороев. Масштаб осенних пороев впечатляет – до 65–80 % на 100 м2. К концу осени интенсивность пороев заметно снижается и достигает всего 15–30 %.
Порои кабана, в результате которых происходит взрыхление и перемешивание почвы, являются хорошим источником гумуса для почвы. Влияние гумусовых веществ, включая гуминовые препараты, на условия питания и развития растений было установлено многими исследователями (Flaig, 1967; Александрова, 1972; Кононова, 1972;). В результате перемешивания поверхностных, гумусовых слоёв почвы, экскрементов кабана и остатков пищи растительного и животного происхождения, почва обогащаеся групповым составом гумуса, в который входят специфические (углерод гумусовых и фульватных кислот) и неспецифические вещества (общий углерод). Динамика группового состава гумуса в разных типах почв и в разных географических широтах выявил неодназначную динамику основных показателей гумусовых соединений в почвах, подверженных механическому воздействия кабана.
Известно, что гуминовые кислоты практически не растворяются в воде и минеральных кислотах и состоят из углерода, водорода, кислорода, азота, серы, фосфора. Также в препаратах гуминовых кислот обнаружены следы Mg, Mn, Na и K, окислов Al и Fe. Содержание этих элементов в гуминовых кислотах колеблется в зависимости от типа почвы, химического состава разлагающихся остатков, условий гумификации и способа выделения (особенность методики определения) (Орлов, 1990). Обогащение почвы вышеперечисленными веществами способствует созданию благоприятных условий для роста и развития растительности и микроорганизмов в почве. Фульвокислоты имеют противоположную направленность своего влияния на почвенный микробоценоз и растительность. Так, если в составе гумусовых веществ много фульвокислот, то почва легко обедняется кальцием, магнием, калием и другими основаниями, так как фульвокислоты образуют с ними растворимые соли, мигрирующие из почвы с просачивающейся влагой. Таким образом, следует отметить, что степень разрушительного действия фульвокислот на минералы зависит от количества гуминовых кислот в данной почве: чем меньше в почве гуминовых кислот, тем сильнее действие фульвокислот (Носко и др., 1988).
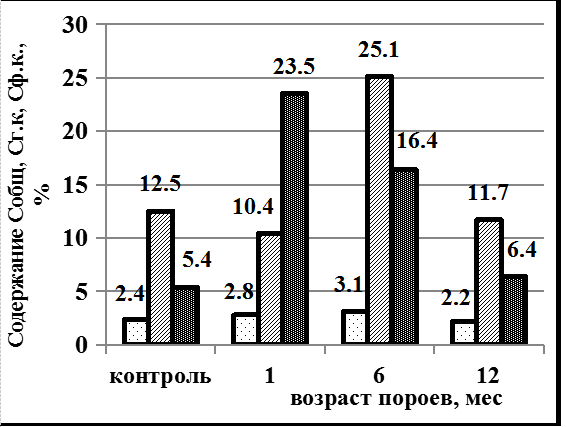
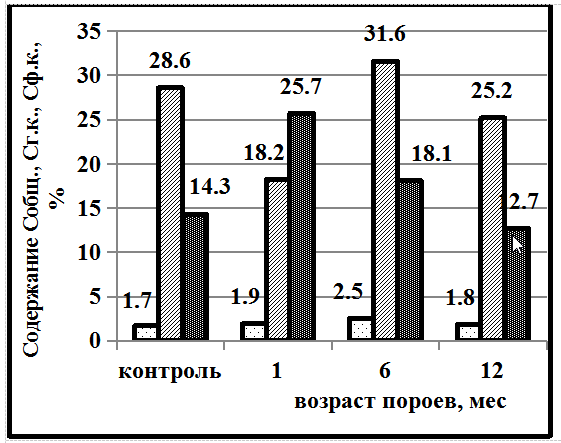
Динамика содержания гуминовых веществ в почвах, подверженных роющей деятельности кабана и без них (контроль) на территории Степного Приднепровья (Украина) приведена на рисунке 1. В черноземной зоне в тяжёлых по гранулометрическому составу почвах – суглинках содержание углерода гуминовых кислот (Сг.к.) значительно выше углерода фульвокислот (Сф.к.), что является положительной тенденцией и связано с характеристикой специфических веществ. В дерново-боровой супесчанной почве содержание углерода гуминовых и фульвокислот практически одинаково.
Под влиянием пороев кабана на обоих типах почв динамика содержания гумусовых веществ идентична. Через 3 месяца наблюдается преобладание Сф.к. над Сг.к. в чернозёме лесном среднесуглинистом в 2,3 раза, в дерново-боровой супесчаной в 1,3 раза. Но уже через 6 месяцев наблюдается обратная тенденция – преобладание содержания гуминовых кислот над углеродом фульвокислот в 1,5 и в 2,4 раза соответственно. Через 12 месяцев показатели гуминовых веществ приближаются к контрольным цифрам (почвы без пороев).
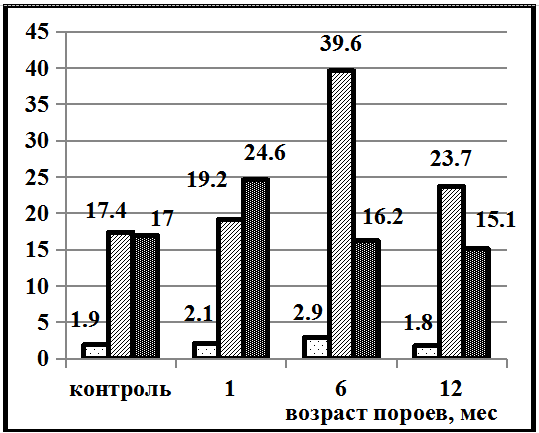
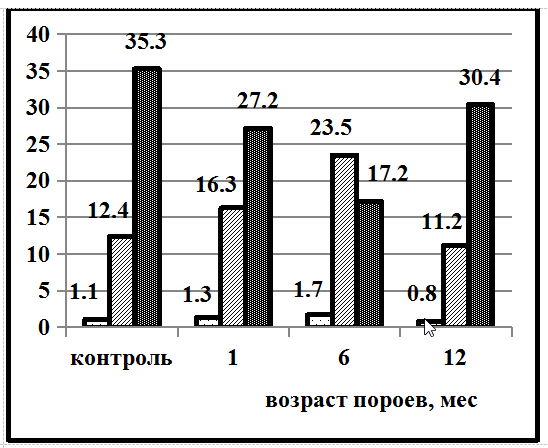
На территории Северо-Запада России (Грязовецкий район Вологодской области) в Ельнике кисличном также прослеживается похожая тенденция в легкосуглинистых почвах (рис. 2).
 |
 |
|
А |
Б |
|
Рис. 1. Почвы степного Приднепровья: А – чернозем лесной среднесуглинистый, Б – дерново-боровая супесчаная почва
|
|
 |
 |
|
А |
Б |
|
Рис. 2. Почвы Грязовецкого района Вологодской области: А – сильноподзолистая легкосуглинистая, Б – сильноподзолистая супесчаная почва
|
|
Но в более лёгких супесчаных почвах динамика содержания гумусовых веществ совершенно иная. В контрольных супесчаных почвах фульвокислоты преобладают над гуминовыми в 2,8 раза. Повышение содержания гуминовых кислот над фульвокислотами наблюдается только через 6 месяцев в 1,4 раза. В остальных случаях зафиксировано преобладание фульвокислот. Данный характер динамики содержания гумусовых веществ в супесчаных почвах можно объяснить повышенной влажностью и кислотностью почв Вологодского региона. Несмотря на высокую растворимость фульвокислот и хорошую промывную способность супесчанных почв, наблюдается повышенное содержание Сф.к. в сильноподзолистой супесчаной почве.
Повышение содержания общего углерода во всех типах почвы через 6 месяцев является типичным явлением и объясняется тем фактом, что в оптимальных условиях, то есть при оптимальной температуре, влажности и времени разложения, полное разложение органического вещества в виде остатков растительной пищи, оставленной кабаном в почве во время кормёжки, а также экскрементов кабана, проходит через 6 месяцев от начала разложения. Общий углерод (Собщ.) – важный почвенный элемент плодородия, необходимый для жизнедеятельности растений и микроорганизмов. Известно положительное влияние Собщ. на тепловой баланс и формирование физико–химических свойств почвы, а также на процессы, протекающие в системе «почва – растение» (Игутинская и др., 1994).
В процессе роюще-рыхлительной деятельности кабана на вырубках после еловых коренных лесов происходит дальнейшая минерализация мёртвых растительных остатков в виде повреждённых корней древесно-кустарниковой растительности. Часть растительного отпада остаётся на пороях, а другая часть в почве. Наиболее насыщены корнями и растительными остатками верхние почвенные горизонты и подстилка, где и происходят основные процессы их отмирания и разложения. Скорость минерализации клетчатки зависит от рельефа, возраста подроста и подлеска, корнями которых питается кабан, глубины залегания грунтовых вод, температурного режима почвы, влажности, от активности микроорганизмов и насекомых, употребляющих органические остатки и так далее. В еловых лесах Вологодской области скорость минерализации тонких корней ели, кустарничков и трав, диаметром около 1 мм, неодинакова (табл. 1).
Таблица 1
Минерализация отмерших корней, выраженная в доле потерь массы за год в еловых коренных лесах Вологодской области
|
Тип ельника, вид опада |
Доля потери массы за год, % |
|
Кисличный |
|
|
Корни ели |
21±2,8 |
|
Корни кустарничков |
32±4,1 |
|
Корни злаков (вейник) |
22±4,7 |
|
Брусничный |
|
|
Корни ели |
17±4,3 |
|
Корни кустарничков |
29±5,2 |
|
Разнотравный |
|
|
Корни ели |
31±3,2 |
|
Корни кустарничков |
42±3,6 |
|
Корни злаков (вейник) |
27±4,7 |
Наиболее быстро разрушаются корни кустарничков (брусника, черника), затем разлагаются мелкие корни молодой ели (до 15 лет), наиболее медленное разрушение выявлено у злаковых (вейник). При этом среднее время разрушения тонких корней (диаметр 1–3 мм) сопоставимо с периодом минерализации листового опада (Глазов, 2004).
ЗАКЛЮЧЕНИЕ
Масштабы средопреобразующей деятельности млекопитающих на эдафотоп довольно значимы. В общем процессе почвообразования она занимает важное место и имеет заметное влияние на всю лесную экосистему. Средопреобразующая деятельность наземных млекопитающих – важный биогеоценотический механизм формирования химических свойств почвы.
Кабан как типичное животное-почворой обеспечивает вертикальную миграцию элементов и соединений в почве, способствуя, тем самым, биологическому круговороту веществ и энергии. Через 6 месяцев почва на месте пороя обогащается гуминовыми кислотами, что способствует некоторому повышению плодородия. Это может вызвать более эффективное прорастание семян, попавших на нарушенное кабаном место в этот период, так как почва не только обогащается полезными специфическими веществами (Сг.к.), но и проходит предварительную минерализацию, то есть сдирание напочвенного покрова с дерниной и взрыхление верхнего слоя.
Таким образом, положительное влияние роющей деятельности на свойства почвы хоть и носит кратковременный и точечный характер, но всё же является неоспоримым вкладом в эффективность существования эдафотопа как сложной живой системы.
Благодарности. Авторы выражают признательность к. б. н., профессору Днепропетровского национального университета имени Олеся Гончара Булахову Валентину Леонтьевичу за консультативную помощь и оригинальные идеи в осуществлении исследований функциональной роли млекопитающих в биогеоценозах.
REFERENCES
Абатуров Б. Д. Деятельность животных-землероев в почвах, ее значение и основные пути изучения // Проблемы почвенной зоологии: Материалы IV Всесоюзного совещания. – М.: Наука, 1972. – С. 5.
Абатуров Б. Д. Млекопитающие как компонент экосистемы. – М.: Наука, 1984. – 286 с.
Александрова И. В. О физиологической активности гумусовых веществ и продуктов метаболизма микроорганизмов // Органическое вещество целинных и освоенных почв. – М.: Наука, 1972. – С. 30–69.
Булахов В. Л. К вопросу о классификации средообразующей деятельности позвоночных животных // Вопросы степного лесоведения: Труды Комплексной экспедиции ДГУ. – 1973. – Вып. 4. – С. 111–116.
Данилов Д. Н., Русаков Л. С., Рыковский А. С., Солдаткин Е. И., Юргенсон Л. Б. – Основы охотустройства. – М.: Лесная промышленность, 1966. – 332 с.
Глазов М. В. Роль животных в экосистемах еловых лесов. – М.: ПАСЬВА, 2004. – 239 с.
Данилкин А. А. Млекопитающие России и сопредельных регионов. Свиные (Suidae). – М.: ГЕОС, 2002. – 309 с.
Злотин Р. И., Исаков Ю. А., Ходашова К. С. Роль животных в функционировании экосистем // Материалы совещания. Московское общество испытателей природы. Институт географии АН СССР. Главное управление охраны природы, заповедников и охотничьего хозяйствава МСХ СССР. – М.: Наука, 1975. – 220 с.
Злотин Р. И., Ходашова К. С. Влияние животных на автотрофный цикл биологического круговорота // Проблемы биогеоценологии. – 1973. – С. 105–117.
Игутинская Г. А., Иванова Н. И., Воцелко С. К., Голобородько С. П. Особенности гумусообразования при сидерации южных орошаемых черноземов // Почвоведение. – 1994. – № 3. – С. 83–89.
Кононова М. М., Бельчикова Н. П. Экспресс-метод определения группового состава гумуса. Опыт характеристики природы почвенных гуминовых кислот с помощью спектрофотометрии // Доклады АН СССР. – Т. 72, № 1. – С. 125–128.
Кононова М. М. Проблема органического вещества на современном этапе // Органическое вещество целинных и освоенных почв. – М.: Наука, 1972. – С. 7–29.
Матюшкин Е. Н. Учеты по следам и территориального размещения зверей в таежных среднегорьях // Экология, методы изучения и организации охраны млекопитающих горных областей. – Свердловск: УНЦ АН СССР, 1977. – С. 94–96.
Новиков Г. А. Полевые исследования по экологии наземных позвоночных. – М.: Изд. Сов. Наука. 1953. – 503 с.
Носко Б. С., Медведев В. В., Бацула А. А. и др. Влияние органических и минеральных удобрений на плодородие почв // Почвы Украины и повышение их плодородия. – Киев: Уроджай, 1988. – Т. 2. – С. 19–34.
Орлов Д. С. Гумусовые кислоты и общая теория гумификации. – М.: Наука, – 1990. – С. 25–32.
ГОСТ 28168-89. Почвы. Отбор проб. – М.: Изд-во стандартов, 1989. – 36 с.
Пахомов А. Е. Биогеоценотическая роль млекопитающих в почвообразовательных процессах степных лесов Украины. – Днепропетровск: ДГУ, 1998. – Т. 1. – 232 с.
Пахомов А. Е. Роющая деятельность млекопитающих как фактор стабилизации лесных почв // Франція та Україна: науково-практичний досвід у контексті діалогу національних культур: тези доп. 5-й Міжн. конф. – Ч. 3. Екологія і медицина. – Дніпропетровськ, 1998а. – С. 13.
Пахомов А. Е., Грицан Ю. И., Курочкина О. Г. Влияние роющей деятельности Sus scrofa L. на изменение температурного режима почв Карадагского заповедника // Екологія та ноосферологія. – 2003. – Т. 13, № 12. – С. 99–103.
Пилипко Е. Н. Влияние роющей деятельности кабана на растительность и напочвенный покров в Устюженском районе Вологодской области // Актуальные вопросы естественных и математических наук в современных условиях развития страны. Сборник научных трудов по итогам международной научно-практической конференции. – Санкт-Петербург, 2015. – № 2. – С. 51–54.
Пилипко Е. Н., Пилипко А. В. Влияние роющей деятельности дикого кабана (S. scrofa L.) на физико-химические показатели почвы в Устюженском районе Вологодской области // Материалы V Международной научно-практической конференции. Фундаментальная наука и технологии – перспективные разработки (24–25 февраля 2015 г.). – North Charleston, USA, – 2015. – Т. 2. – С. 7–9.
Тюрин И. В. Органическое вещество почвы и его роль в плодородии. – М.: Наука, – 1965. – С. 78–97.
Формозов А. Н. Следы животных и методы их «тропления» при изучении наземных животных // Справочник путешественника и краеведа. – М.: Географгиз, 1950. – Т. 2. – С. 536–552.
Формозов А. Н. Спутник следопыта. – М.: Изд-во МОИП, 1952. – 360 с.
Челинцев Н. Г. Методы расчета плотности населения животных по данным маршрутных учетов // В кн. Пространственно-временная динамика животного населения. – Новосибирск: Наука, 1985, – С. 5–14.
Flaig W. Chemical composition and physical properties of humic substances // Studies about gumus: Transact. of the Intern. symp. «Humus et planta, IV”. – Prague, 1967. – Р. 81–112.
