
WATER REGIME OF SOME REPRESENTATIVES OF THE GENUS IRIS L. DURING INTRODUCTION IN THE SOUTHERN URALS
JOURNAL: «EKOSISTEMY», Issue 22, 2020
Publication text (PDF):Download
UDK: 582.579.2:581.111:581.522.4
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
- Beksheneva L. F., South-Ural Botanical Garden-Institute of the Ufa Federal Research Centre of the Russian Academy of Sciences, Ufa, Republic of Bashkortostan, Russia
- Reut A. A., South-Ural Botanical Garden-Institute of the Ufa Federal Research Centre of the Russian Academy of Sciences, Ufa, Republic of Bashkortostan, Russia
TYPE: Article
DOI:https://doi.org/10.37279/2414-4738-2020-22-82-89
PAGES: from 82 to 89
STATUS: Published
LANGUAGE: Russian
KEYWORDS: Iris, water regime, hydration, water retention capacity, water deficit, botany, Republic of Bashkortostan.
ABSTRACT (ENGLISH):
The article presents the results of a study of the parameters of the water regime of irises of the subgenus Xyridion introduced in the Southern Urals (Iris graminea, I. halophila, I. halophila var. sogdiana, I. notha, I. orientalis, I. spuria, I. spuria subsp. carthaliniae, I. spuria subsp. musulmanica) in comparison with local species (I. sibirica, I. pseudacorus, I. pumila). The experiments were carried out on the basis of the laboratory of introduction and selection of flower plants of the South-Ural Botanical Garden-Institute of Ufa Federal Research Centre of Russian Academy of Sciences. The objects of the study were different ecological groups of plants in relation to water ‒ hydrophytes, mesophytes and xeromesophytes. The water deficit of leaves and accompanying characteristics (relative turgescence) were determined by saturation of plant samples according to methodological guidelines; water retention capacity, tissue hydration and the level of daily water loss were determined by artificial wilting. The obtained results characterize the peculiarities of the water regime of irises and indicate significant interspecific differences. It was revealed that the flora of the Republic of Bashkortostan contains plants both with a wide range of variability of water retention capacity (I. pumila) and with a fairly conservative one (I. pseudacorus, I. sibirica). Both strategies are successful in the continental climate of the Southern Urals. Introduced taxa of the subgenus Xyridion also keep to a particular strategy and it can be considered an indicator of successful introduction. It is shown that local species of irises do not form a water deficit in conditions of excessive moisture, while 6 taxa of the subgenus Xyridion have an average level of water deficit. According to the sum of indicators, the studied taxa belong to the groups with medium and high stability of the water regime due to low water deficit, high water retention capacity and hydration. These parameters are found in various combinations and sustain the stability of the water regime of cultivars. The studied physiological indicators can be used to assess the success of introduction in comparison with local control species.
Виды ирисов освоили широкий диапазон местообитаний – от прибрежных до пустынных. Среди них нет настоящих лесных растений, они предпочитают открытые пространства и прогреваемые участки. Среди ирисов есть псаммофиты, отличающиеся высокой жаро- и засухоустойчивостью, гидрофиты, кальцефобы, виды, способные переносить высокую засоленность грунта, и виды, отличающиеся исключительно высокой холодостойкостью (Алексеева, 2008). В коллекции иридария Южно-Уральского ботанического сада-института ‒ обособленного структурного подразделения Федерального государственного бюджетного научного учреждения Уфимского федерального исследовательского центра РАН (далее ЮУБСИ УФИЦ РАН) собраны ирисы из различных регионов и климатических поясов, в том числе виды подрода Xyridion, не затрагивающие своим ареалом Южное Предуралье. Процесс адаптации (акклиматизации) растений к определенным условиям сопровождается активным приспособлением организма к непривычным для него условиям (Елисафенко, 2009). Среди физиологических показателей приспособленности растений к данным местным условиям одно из важных мест занимает водный режим растений, включающий в себя ряд параметров. Их одновременный учет и изучение в конкретных условиях позволяют дать оценку адаптационным возможностям и засухоустойчивости растений (Миронова и др., 2010).
Водоудерживающую силу клеток обеспечивают осмотически активные и коллоидные вещества. В регуляции водообмена водоудерживающая способность зависит также от реакции устьичного аппарата на воздействие экстремальных условий (Голяева, Петров, 2007). Растения, устойчивые к засухе, отличаются высокой водоудерживающей способностью, то есть отдают в процессе завядания меньше воды, чем листья менее устойчивых (Жолкевич и др., 1989). Водоудерживающая способность растений определяется количеством потерянной воды в процессе обезвоживания листьев за более продолжительный, чем при определении транспирации, отрезок времени.
Оводненность тканей органов – также важный показатель водного режима растений. В засушливых условиях уровень общей оводненности тканей снижается. Однако этот показатель не объясняет, находится ли растение в состоянии водного насыщения или испытывает его недостаток. Степень насыщения тканей водой характеризуется величинами водного дефицита и относительной тургесцентности. В благоприятных условиях или при избыточном увлажнении водный дефицит имеет низкие величины и возрастает в условиях недостаточного увлажнения (Самуилов, Мухитов, 2012).
Водный дефицит образуется, когда поступление воды в растения не успевает за ее расходованием. В нормальных условиях водоснабжения водный дефицит компенсируется возрастанием сосущей силы листьев. При недостатке водоснабжения может наблюдаться завядание растений. У некоторых видов тенистых местообитаний потеря воды, равная 3‒5 %, уже вызывает завядание. Однако у других видов завядание наступает при водном дефиците 20‒30 % (Якушкина, 1980).
Считается, что полная насыщенность растительных тканей влагой не является лучшей для роста и развития растений. Оптимальная оводненность характеризуется небольшим водным дефицитом (Миронова и др., 2014). В частности, для пшеницы он составляет 3‒10 % (Жолкевич и др., 1989). Общепринято, что водный дефицит, не превышающий 10 %, не причиняет растению вреда. При 25 %-ном водном дефиците происходит завядание листьев, закрывание устьиц, снижение интенсивности фотосинтеза и роста, нарушение синтеза в клетках (Макараев, Титова, 2003).
Цель настоящего исследования – изучить некоторые особенности водного режима ирисов подрода Xyridion, интродуцированных в Южное Предуралье, в сравнении с местными видами.
МАТЕРИАЛ И МЕТОДЫ
Исследования проводились на базе лаборатории интродукции и селекции цветочных растений ЮУБСИ УФИЦ РАН. Почвы на опытном участке серые лесные, типичные для региона, pH=5,89.
По метеоданным нами был рассчитан гидротермический коэффициент Г. Т. Селянинова (Чирков, 1986), который является показателем обеспечения территории влагой (табл. 1). Характеристики рассчитывались по материалам архива погоды сайта http://rp5.ru/. Образцы листьев для анализа отбирали дважды, в июле и августе 2019 года. Все взвешивания производили на электронных лабораторных весах Госмер ВЛТЭ 1100.
Таблица 1
Характеристика влагообеспеченности вегетационного периода 2019 года
| Гидротермический коэффициент по месяцам | ГТК за год | Характеристика влагообеспеченности года | ||||||
| апрель | май | июнь | июль | август | сентябрь | октябрь | ||
| 0,9 | 2,3 | 1,4 | 1,2 | 2,5 | 1,8 | 1,4 | 1,8 | Избыточно увлажненный |
Примечание к таблице. Градации по Г. Т. Селянинову ‒ зона избыточного увлажнения, или зона дренажа (ГТК>1,3), зона обеспеченного увлажнения (1,0–1,3); зона засушливая (0,7–1,0), зона сухого земледелия (0,5–0,7), зона сухая или ирригации (ГТК<0,5).
В работе были использованы следующие методы для определения основных показателей водного режима. Водный дефицит листьев и сопутствующие характеристики (относительная тургесцентность) определяли методом насыщения растительных образцов по методическим указаниям (Макараев, Титова, 2003); водоудерживающую способность, оводненность тканей, уровень суточной водопотери ‒ методом искусственного завядания (Гусев, 1960, Таренков, Иванова, 1990). Опыты ставили параллельно. Листья собирали в период с 10 до 11 часов, измерения проводились с интервалом в месяц.
Объекты исследования – 11 таксонов (8 видов и 3 подвида) рода Iris (рис. 1). 
Рис. 1. Некоторые объекты исследования
a – Iris graminea; b – I. halophila; c – I. orientalis; d – I. spuria.
Таксоны подрода Xyridion (8 штук) не произрастают в Южном Предуралье и все являются интродуцированными. В группах Limniris и Iris мы выделили эталонные маркерные виды, обитающие в Республике Башкортостан: I. sibirica, I. pseudacorus, I. pumila. Данные виды представляют собой различные экологические группы растений по отношению к воде – гидрофиты, мезофиты, ксеромезофиты (табл. 2). Экологические характеристики видов приведены согласно литературным источникам (Алексеева, 2008, Кирпичева, 2009, Шевченко, Скрипчинская, 2009, Бородич, 2012, Nadiradze, Eradze, 2014, Бирюкова и др., 2018).
Таблица 2
Объекты исследования и их экологическая характеристика
| Виды | Природный ареал | Экологическая группа |
| Подрод Xyridion | ||
| Iris graminea | Европа до Украины, Кавказ | Ксеромезофит, полуксерофит, условно-водолюбивый |
| Iris halophila | Восточная Европа, Кавказ, юг Европейской части РФ и Западной Сибири, Средняя и Центральная Азия | Гидрофит, ксеромезофит |
| Iris halophila var. sogdiana | Средняя Азия, умеренные регионы Центральной Азии | Гидрофит |
| Iris notha | Предкавказье, Ростовская область | Мезофит, сухолюб |
| Iris orientalis | Турция, юго-восточная Европа (Греция) | Ксеромезофит, условно-водолюбивый, влаголюбив |
| Iris spuria | Европа, Кавказ, Сибирь, Китай, Средняя Азия, Индия, Пакистан, Северная Африка | Мезофит |
| Iris spuria subsp. carthaliniae | Кавказ, Турция | Мезофит, гидрофит, условно-водолюбивый |
| Iris spuria subsp. musulmanica | Кавказ, Турция, Иран | Гидрофит |
| Подрод Limniris | ||
| Iris pseudacorus | Африка, Европа, Кавказ, Западная Азия, Западная Сибирь | Гидрофит |
| Iris sibirica | Восточная и Центральная Европа, Северная Азия | Гидрофит, мезофит |
| Подрод Iris | ||
| Iris pumila | Степная зона Европы | Ксеромезофит, полуксерофит |
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Для ранжировки сортов по засухоустойчивости существуют шкалы, разработанные для сельскохозяйственных культур (Гончарова, 1988). Опираясь на полученные результаты, мы разработали подобную шкалу для ирисов (табл. 3).
Таблица 3
Устойчивость водного режима растений рода Iris
| Параметры | Степень устойчивости | ||
| высокая | средняя | низкая | |
| Оводненность тканей, % | ≥80 | 60‒79 | ≤59 |
| Водоудерживающая способность, % | ≥50 | 30‒49 | ≤29 |
| Водный дефицит, % | 0‒15 | 16‒29 | ≥30 |
В целом, вегетационный период 2019 года можно отнести к категории «избыточно увлажненный» (ГТК=1,8). Июль 2019 года характеризовался достаточно увлажненной погодой ‒ ГТК составил 1,2; максимальная температура достигала 31,3 °С при 39 мм осадков (табл. 1). В этом месяце таксоны подрода Xyridion имели средние значения общей оводненности (66,84‒75,13 %), за исключением I. spuria – чуть ниже средних значений (57,02 %). Значения показателя для листьев контрольных видов из различных экологических групп оказались практически одинаковыми – 74,59‒6,74 %. Пять таксонов подрода Xyridion ‒ I. graminea, I. halophila, I. halophila var. sogdiana, I. orientalis, I. spuria subsp. carthaliniae – наиболее близки по значениям параметра к группе местных видов. I. spuria, значительно уступающий контрольным видам, относится к мезофитам. На 6 и 8 % соответственно показатель меньше у I. notha (мезофит) и I. spuria subsp. musulmanica (гидрофит).
Август 2019 года оказался избыточно увлажненным – ГТК равен 2,5 при 76 мм осадков. Максимальная температура достигала 30,8 °С. Логично предположить, что в условиях избыточного увлажнения показатели общей оводненности увеличатся. Однако в августе значение параметра в группе местных видов увеличилось только для I. pumila ‒ на 12 % (92,31 %), осталось таким же для I. pseudacorus (72,69 %) и уменьшилось на 10 % для I. sibirica (65,02 %).
При таких погодных условиях для ирисов подрода Xyridion значения параметра в целом оказались более высокими по сравнению с июлем ‒ от 64,53 до 84,77 %. В исследованной группе выделяются виды с высоким уровнем оводненности ‒ I. graminea и I. halophila var. sogdiana. Остальные таксоны характеризуются средними значениями параметра. Таким образом, в условиях достаточного и избыточного увлажнения тканям ирисов подрода Xyridion свойственна средняя или высокая оводненность.
Рассматривая группу Xyridion в свете экологических характеристик, можно заметить что, например, ксеромезофиты I. graminea и I. orientalis в данном случае имеют разные стратегии в условиях избыточного увлажнения: первый увеличивает свою оводненность и этим близок к I. pumila, а второй – уменьшает, приближаясь по значению параметра к I. sibirica. I. halophila, который разными исследователями определяется как в группу гидрофитов, так и в группу ксеромезофитов (табл. 2), несмотря на избыточное увлажнение, также снижает оводненность до уровня I. sibirica.
Общая оводненность гидрофитов (I. halophila var. sogdiana, I. spuria subsp. musulmanica) в августе отличается от таковой у I. pseudacorus, в ту и другую сторону. I. halophila var. sogdiana увеличивает оводненность с 75,13 до 84,77 %, в то же время значение показателя остается стабильным для I. spuria subsp. musulmanica в оба месяца (66,84‒66,33 %). Стабильность показателя по месяцам также характерна для I. notha (мезофит).
Подводя итог проведенному анализу, мы можем заключить, что содержание воды в органах растений разных экологических групп не различается в условиях «достаточного увлажнения». В условиях «избыточного увлажнения» также не наблюдается никакой зависимости общей оводненности от принадлежности таксона к определенной экологической группе.
По показателю общей оводненности невозможно полно оценить способность растений к регуляции водного обмена (его устойчивость), поэтому в рамках исследования изучалась водоудерживающая способность после суточного завядания, а также водный дефицит. Данные показатели также были ранжированы на три группы (табл. 3).
В июле I. notha имеет самое низкое значение водоудерживающей способности – всего 11,87 %. На другой стороне интервала ‒ I. graminea, показатель равен 49 %. Остальные таксоны укладываются в довольно узкий промежуток между 27,19‒37,21 %, попадая в группу низких или средних значений параметра, в том числе контрольные виды ‒ гидрофит I. pseudacorus и ксеромезофит I. pumila. Другой маркерный вид, мезофит I. sibirica, имеет низкую водоудерживающую способность – 21,79 % (рис. 1).
В августе для I. graminea отмечено резкое снижение водоудерживающей способности (до 29,79 %) при сохранении высокой оводненности. На 8,7 % снижается значение параметра у I. spuria subsp. carthaliniae. Остальные таксоны, независимо от принадлежности к экологической группе, в условиях высокой увлажненности демонстрируют рост показателя, порой довольно интенсивный. Так, значение параметра увеличилось на 31,01‒39,71‒41,98 % для I. halophila var. sogdiana, I. pumilа, и I. notha и составило соответственно 60,92–76,92–
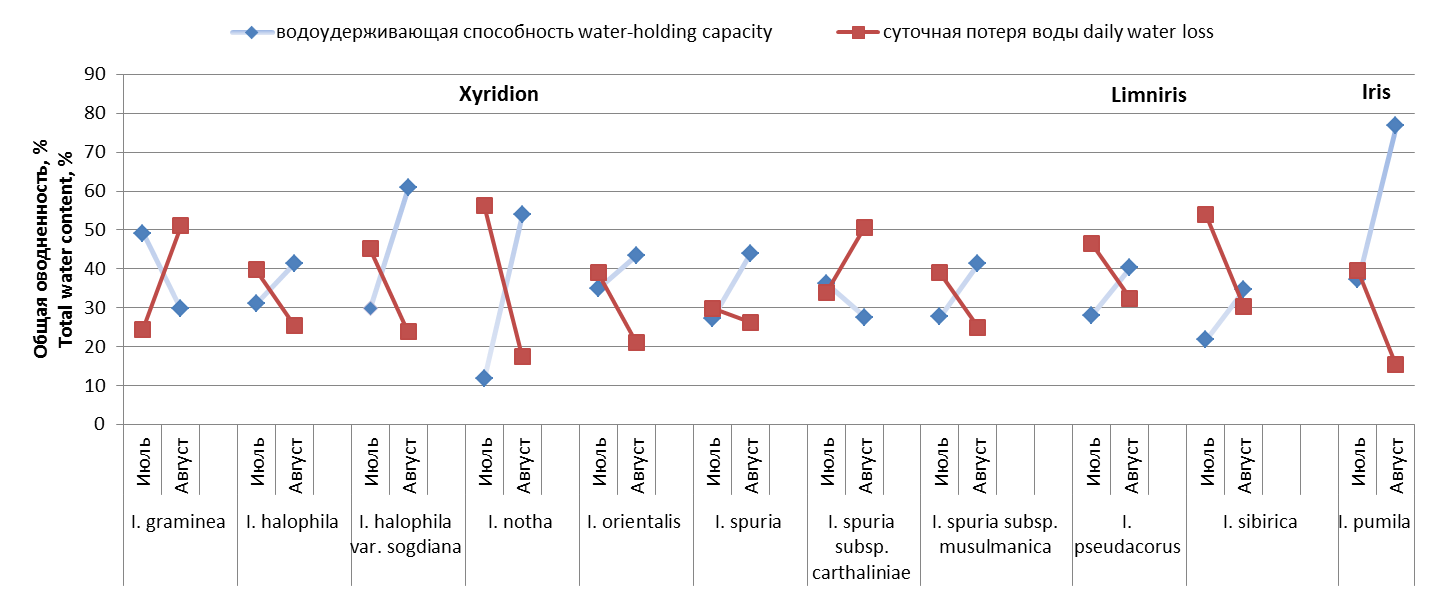
Рис. 1. Показатели водного режима некоторых видов ирисов
53,95 %. Контрольные виды ‒ I. pseudacorus, I. sibirica демонстрируют рост параметра на 12,16‒12,96 %. Для 4 таксонов подрода Xyridion ‒ I. halophila, I. orientalis, I. spuria, I. spuria subsp. musulmanica – рост составил 8,52‒16,73 % (рис. 1).
Таким образом, во флоре РБ присутствуют виды как с широким диапазоном изменчивости водоудерживающей способности (I. pumilа), так и с довольно консервативным (I. pseudacorus, I. sibirica). Обе стратегии успешны в условиях континентального климата Южного Предуралья. Интродуцированные таксоны подрода Xyridion также придерживаются той или иной стратегии, что может служить показателем успешной интродукции.
Показатели водного дефицита определялись в августе, в условиях избыточной водоопеспеченности (табл. 4).
В дни измерений водного дефицита относительная влажность воздуха достигала 80 %. Ожидалось, что в подобных условиях дефицит водного насыщения будет сведен к минимуму как у местных, так и у интродуцированных видов. Cреди исследованных видов водный дефицит не сформировался (0 %) у местного вида I. pumilа. Низкий водный дефицит от 5 до 12 % отмечен для других контрольных видов ‒ I. sibirica и I. pseudacorus, а также для I. notha и I. spuria subsp. сarthaliniae. Некоторые авторы считают, что такие величины не говорят о наличии водного дефицита (Евдокимова и др., 2012). Для остальных таксонов подрода Xyridion величина показателя составила 15,08‒5,78 %. Наибольший дефицит воды имеют обитатели влажных местообитаний ‒ I. halophila var. sogdiana, I. orientalis – выше 20 %. При этом данные виды имеют высокие или средние показатели водоудерживающей способности, что позволяет отнести их к видам со стабильным водным режимом.
Таким образом, ожидаемое в условиях избыточного увлажнения отсутствие или минимальное наличие водного дефицита подтвердилось на практике для местных и 2-х интродуцированных таксонов подрода Xyridion. Наличие водного дефицита среднего уровня в условиях отсутствия водного стресса отмечено для 6 таксонов подрода Xyridion.
Мы не рассматриваем в данной статье суточную динамику водного дефицита. Так, во влажный сезон в тропическом лесу для разных видов разница между средними и максимальными значениями водного дефицита может составлять от 1,4 до 16 %, достигая максимума в дневное время и минимума в утренние и вечерние часы (Евдокимова и др., 2012). В нашем случае, одномоментное измерение параметра позволяет провести сравнение интродуцированных видов с контрольными. Но мы также можем предположить возможные различия в суточной динамике водного дефицита, в частности наличие пика водного дефицита у интродуцированных растений, не совпадающего с максимумом местных видов.
Таблица 4
Водный дефицит листьев растений рода Iris в августе
| Виды | Водный дефицит, % | Относительная тургесцентность, % |
| Подрод Xyridion | ||
| Iris graminea | 15,46 | 84,54 |
| Iris halophila | 18,25 | 81,75 |
| Iris halophila var. sogdiana | 25,78 | 74,22 |
| Iris notha | 10,84 | 89,16 |
| Iris orientalis | 20,46 | 6,46 |
| Iris spuria | 16,79 | 83,2 |
| Iris spuria subsp. carthaliniae | 5,71 | 94,29 |
| Iris spuria subsp. musulmanica | 15,08 | 84,92 |
| Подрод Limniris | ||
| Iris pseudacorus | 12,91 | 87,09 |
| Iris sibirica | 5,49 | 94,51 |
| Подрод Iris | ||
| Iris pumila | 0 | 100 |
Относительная тургесцентность – обратный водному дефициту показатель: чем больше водный дефицит, тем меньше доля воды, участвующей в обеспечении тургора растительных тканей. В соответствии с этим значения относительной тургесцентности максимальны для ирисов с низким водным дефицитом (табл. 4).
ЗАКЛЮЧЕНИЕ
Полученные результаты характеризуют особенности водного режима ирисов подрода Xyridion и свидетельствуют о следующих межвидовых различиях. Согласно проведенным исследованиям, содержание воды в ирисах не зависит от принадлежности таксона к определенной экологической группе. Среди контрольных и интродуцированных видов имеются таксоны, как с широким, так и с узким диапазоном изменчивости водоудерживающей способности. Местные виды ирисов в условиях избыточного увлажнения не формируют водный дефицит. Большая часть видов подрода Xyridion в тех же условиях характеризуется средним водным дефицитом, что, возможно, связано с суточной динамикой параметра.
В целом, по сумме показателей исследованные виды ирисов относятся в группы со средней и высокой устойчивостью водного режима за счет низкого или среднего водного дефицита, средней или высокой оводненности и водоудерживающей способности. Эти параметры встречаются в различных сочетаниях и обеспечивают стабильность водного режима касатиков. Показатели водного режима вполне подтверждают успешность интродукции ирисов подрода Xyridion в условиях Южного Предуралья.
Работа выполнена по Программе фундаментальных исследований Президиума РАН «Биоразнообразие природных систем и биологические ресурсы России» и в рамках государственного задания ЮУБСИ УФИЦ РАН по теме АААА-А18-118011990151-7
REFERENCES
Алексеева Н. Б. Род Iris L. (Iridaceae) в России // Turczaninowia. ‒ 2008. ‒ Т. 11, № 2. ‒ С. 5‒70.
Бирюкова О. А., Куклина Е. Э., Гринченко К. В., Зайцева Д. М., Куршакова Е. И. Показатели плодов ирисов подрода Ксиридион в условиях республики Марий Эл // Международный журнал гуманитарных и естественных наук. ‒ 2018. ‒ № 10–1. ‒ С. 121‒124.
Бородич Г. С. Виды и сорта ирисов (Iris) в Центральном ботаническом саду НАН Беларуси // Весці Нацыянальнай акадэміі навук Беларусі. Серыя біялагічных навук. ‒ 2012. ‒ № 1. ‒ С. 22‒25.
Голяева О. Д., Петров А. В. Засухоустойчивость сортов красной смородины // Селекция и сорторазведение садовых культур (сб. научн. тр.). ‒ Орел: Всеросийский научно-исследовательский институт селекции плодовых культур, 2007. ‒ С. 64‒73.
Гончарова Э. А. Оценка устойчивости к разным стрессам плодово-ягодных и овощных (сочноплодных) культур // Диагностика устойчивости растений к стрессовым воздействиям: методическое руководство / [Ред. д. б. н., проф. Г. В. Удовенко]. ‒ Л.: ВИР, 1988. ‒ С. 46‒62.
Гусев Н. А. Некоторые методы исследования водного режима растений. ‒ Л.: АН СССР, Всесоюзное ботаническое общество, 1960. ‒ 60 с.
Евдокимова Е. В., Новичонок А. О., Марковская Е. Ф., Курбатова Ю. А. Особенности водного режима некоторых видов растений в тропическом лесу на юге Вьетнама во влажный сезон // Ученые записки Петрозаводского государственного университета. ‒ 2012. ‒ № 4 (125). ‒ С. 19‒24.
Елисафенко Т. В. Оценка результатов интродукционной работы на примере редких видов сибирской флоры // Растительный мир Азиатской России. ‒ 2009. ‒ № 2 (4). ‒ С. 89‒95.
Жолкевич В. Н., Гусев Н. А., Капля А. В. и др. Водный обмен растений. ‒ М.: Наука, 1989. ‒ 256 с.
Кирпичева Л. Ф. Генофонд ирисов ботанического сада Таврического национального университета им. В. И. Вернадского // Бюллетень Государственного Никитского ботанического сада. ‒ 2009. ‒ № 99. ‒ С. 24‒25.
Миронова Л. Н., Реут А. А., Денисова С. Г., Биглова А. Р., Аллаярова И. Н. Сравнительный анализ адаптационного потенциала декоративных травянистых многолетников // Субтропическое и декоративное садоводство. ‒ 2014. ‒ Вып. 50. ‒ С. 239‒244.
Миронова Л. Н., Реут А. А., Денисова С. Г., Зайнетдинова Г. С., Шайбаков А. Ф., Биглова А. Р., Аллаярова И. Н. Сравнительный анализ жаростойкости и водного режима декоративных травянистых многолетников // Вестник Башкирского университета. ‒ 2010. ‒ Т. 15, № 4. ‒ С. 1153‒1154.
Самуилов Ф. Д., Мухитов Л. А. Водный режим и водопотребление сортов яровой мягкой пшеницы различных экологических групп в контрастных условиях водообеспеченности // Доклады Российской академии сельскохозяйственных наук. ‒ 2012. ‒ № 5. ‒ С. 10‒13.
Таренков В. А., Иванова Л. Н. Водоудерживающая способность листьев боярышников в связи с устойчивостью к засухе // Интродукция, акклиматизация, охрана и использование растений (сб. научн. тр.). ‒ Куйбышев: Куйбышевский госуниверситет, 1990. ‒ С. 3‒9.
Чирков Ю. И. Агрометеорология. ‒ Л.: Гидрометеоиздат, 1986. ‒ 296 с.
Шевченко Г. Т., Скрипчинская Е. А. Виды и культивары Спуриа ирисов как этап интродукции родового комплекса Iris L. в Центральном Предкавказье // Субтропическое и декоративное садоводство. ‒ 2009. ‒ № 42‒1. ‒ С. 202‒206.
Маракаев О. А., Титова О. В. Экологическая физиология растений: вегетационные опыты. ‒ Ярославль: Ярославский государственный университет, 2003. ‒ 55 с.
Якушкина Н. И. Физиология растений. ‒ М.: Просвещение, 1980. ‒ 303 с.
Nadiradze T., Eradze N. In situ conservation of some rare and endemic species of Iridaceae family in National botanical garden of Georgia // European researcher. ‒ 2014. ‒ N 6–2 (77). ‒ P. 1117‒1121.
