
Экосистемы, 29: 5–26 (2022) https://ekosystems.cfuv.ru
![]()
Publication text (PDF): Download
УДК 57.04:574.9:502.4(470.620)
Хорологические закономерности в экотоне Восточно-Средиземноморской и Колхидской биогеографических провинций на территории Сочинского национального парка
Туниев Б. С.1, Рыбак Е. А.2, 1, Тимухин И. Н.1, Алиев Х. У.3, 1
- Сочинский национальный парк Сочи, Россия btuniyev@mail.ru;timukhin77@mail.ru
- Субтропический научный центр РАН Сочи, Россия elena.rybak@gmail.com
- Горный ботанический сад Дагестанского федерального исследовательского центра РАН Махачкала, Россия
Приведен обзор 11 точек исследования биоты и микроклимата в предгорной полосе территории Сочинского национального парка (СНП). В дополнение к субсредиземноморским прибрежным ландшафтам, в СНП на расстоянии до 10 км от моря прослежена цепочка территорий, контрастирующих с окружающими фитоландшафтами, с высокой представленностью средиземноморских видов. Непосредственный контакт биот разного состава и происхождения, их мозаичное распространение на удалении от морского побережья ставит закономерный вопрос о причинах формирования и сохранения экотона двух биогеографических провинций на относительно ограниченной территории СНП. Целью исследований явилась попытка определить особенности развития восточно-средиземноморской и колхидской биот в экотоне двух одноименных биогеографических хорионов на территории СНП с позиций микроклиматических характеристик различных фито-ландшафтов. На формирование экотона двух биогеографических провинций на территории СНП оказывают определенное влияние изменения по количеству осадков, значениям минимальных (средних и абсолютных) температур, средняя амплитуда относительной влажности воздуха, зона воздействия вторжения зимних холодных северо-восточных воздушных масс (бора), в свою очередь определяемые гипсометрической высотой Главного Кавказского хребта, резко понижающегося к западу от поселка Лазаревское. Исследование показало, что для средиземноморских видов приоритетное значение имеет влажность воздуха. В свете приведенных фактов изменения климатических показателей и кружева ареалов колхидских и восточно-средиземноморских видов флоры и фауны авторы предполагают, что в дальнейшем произойдет смещение к востоку условной линии, разделяющей Колхидскую и Восточно-Средиземноморскую биогеографические провинции.
Ключевые слова: Колхида, Восточное Средиземноморье, экотон, Сочинский национальный парк, хорология, микроклиматические особенности.
ВВЕДЕНИЕ
Территория Сочинского национального парка (СНП), площадью 208 599.85 га (~2086 км2), расположена между 40°30’44°15′ с. ш. и 43°30’44°05′ в. д. от Гринвича между рекой Магри на северо-западе и рекой Псоу на юго-востоке (рис. 1). Одной из определяющих особенностей территории СНП является прохождение здесь границы биохорионов высокого порядка. Подавляющая часть территории СНП относится к Колхидской биогеографической провинции, северо-западная оконечность парка входит в состав Восточно-Средиземноморской провинции. Граница между биохорионами на территории СНП проходит от берега Черного моря по водоразделу рек Аше и Псезуапсе до гребня Главного хребта, а затем продолжается за пределами СНП (Tuniyev, 1997).
ISSN 2414-4738 Published by V. I. Vernadsky Crimean Federal University, Simferopol
Туниев Б. С., Рыбак Е. А., Тимухин И. Н., Алиев Х. У.
![]()
При неизменной северной границе Колхиды, проводимой по Главному Кавказскому хребту, западную границу различные авторы проводили (с запада на восток) по реке Джубга (Зернов, 2000), по реке Шапсухо (Зернов, 2006), по водоразделу между реками Шапсухо и Нечепсухо (Соколов, 1931), по реке Туапсе (Меницкий, 1991), южнее Туапсе (Сатунин, 1912). Западные границы Колхиды, проведенные вышеперечисленными авторами севернее междуречья Псезуапсе – Аше, базировались на границах ареалов отдельных мезофильных широколиственных пород и, в первую очередь, Castanea sativa Miller. Однако, если брать за основу секторальный многопоясный подход, очевидна северо-западная граница развития полночленных колонок высотных поясов, характерных для Колхиды, от бассейна реки Псезуапсе и восточнее. Здесь, в истоках реки Псезуапсе на горе Аутль расположены наиболее западные субальпийские луга и расположены западные пределы ареалов для большинства высокогорных видов на Большом Кавказе (Тимухин, Туниев, 2016; Тиимухин, 2019). Исходя из этих особенностей, западная граница Колхиды и флористического Туапсе-Адлерского района по Ю. Л. Меницкому (1991), или Сочинского подрайона Севро-Колхидского района (Тимухин, Туниев, 2016), нами проведена по водоразделу рек Аше и Псезуапсе (рис. 1), соответственно увеличивая протяженность на восток Пшадско-Джубгинского (Меницкий, 1991), или Пшадско-Туапсинского (Тимухин, Туниев, 2016) района Северо-Западного Закавказья. При этом Бело-Лабинский и Туапсе-Адлерский (Сочинский) районы могут рассматриваться не более, чем подрайонами единого Северо-Колхидского района Западного Закавказья.
- приморской полосе СНП эксклавами сохраняется субсредиземноморская растительность, особенно ярко представленная в Макопсинском, Лыготхском и Лазаревском участковых лесничествах. В этих же лесничествах достигает максимального развития пояс гемиксерофильных дубняков и грабинниковых шибляков при резком сокращении верхне-лесных поясов букняков и, особенно, пихтарников.
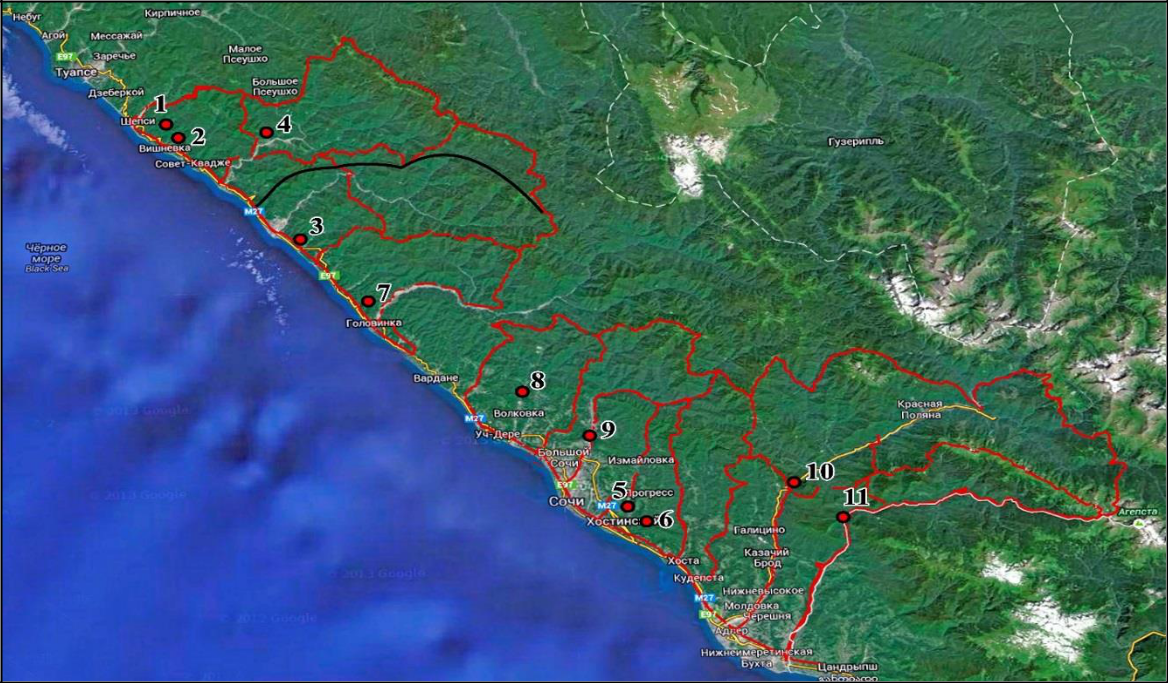
Рис. 1. Границы Сочинского национального парка и точки изучения биоты и микроклимата
Условные обозначения: красные линии – границы участковых лесничеств СНП, черная линия – граница Колхидской и Восточно-Средиземноморской биогеографических провинций. Названия локалитетов приведены в таблице 1 в соответствии с нумерацией.
6
Хорологические закономерности в экотоне Восточно-Средиземноморской и Колхидской биогеографических провинций на территории Сочинского национального парка
![]()
Помимо субсредиземноморских приморских ландшафтов, в СНП, в удалении до 10 км от моря прослеживается цепочка участков, контрастирующих с окружающими фито-ландшафтами высоким представительством средиземноморских видов. В пределах СНП с запада на восток – это вершины Большой Псеушхо – Хакукай – Бозтепе – ущелье среднего течения реки Чимит (Зубова Щель) – ущелье среднего течения реки Западный Дагомыс – Орлиные скалы – гора Большой Ахун – окрестности поселка Веселое (Туниев и др., 2014).
Непосредственный контакт различных по составу и происхождению биот, их мозаичное располажение на удалении от морского берега вызывает закономерный вопрос о причинах формирования и сохранения экотона двух биогеографических провинций на сравнительно ограниченной территории СНП.
Цель исследования – попытка определить особенности развития восточно-средиземноморской и колхидской биот в экотоне двух одноименных биогеографических хорионов на территории СНП, с позиций микроклиматических характеристик различных фито-ландшафтов.
МАТЕРИАЛ И МЕТОДЫ
Материал собран по общепринятым методикам флористических и фаунистических исследований в период 1998–2020 годов, в ходе экспедиционных, стационарных и полустационарных исследований в СНП на отрезке Черноморского побережья, протяженностью около 105 км от междуречья рек Магри и Шесси до государственной границы РФ и Республики Абхазия – реки Псоу в приморской и предгорной зонах в диапазоне высот от 0 до 600 м н. у. м. Основой фиксации флористической информации явились гербарные сборы, которые хранятся в научном отделе СНП (зарегистрирован в Index Herbariorum – SNP). Ботаническая номенклатура дается по «Конспекту флоры Кавказа» (2003, 2006, 2008, 2012), в случае отсутствия в Конспекте, – по С. К. Черепанову (1995) и А. С. Зернову (2000, 2006).
Основой герпетофаунистических исследований являлся маршрутный метод, охватывающий однородные фитоландшафты по всему предгорному поясу и, конкретно, в районах изучения микроклимата. На маршрутах и площадках фиксировались встречи всех представителей амфибий и рептилий. Биогеографические группы амфибий и рептилий даны по классификации Б. С. Туниева (1990).
Изучение микроклимата проводили с использованием автоматических датчиков климата (комплекс измерений IBDL с регистрацией температуры и влажности DS1923-F5) в 11 точках,
- том числе 6 в средиземноморских фито-ландшафтах: приморских – 3 (окрестности поселка Вишнёвка, река Неожиданная, окрестности поселка Солоники), удаленных локальных – 3 (г. Хакукай, Орлиные скалы, г. Большой Ахун); 5 в колхидских фито-ландшафтах: на западной границе хориона – 2 (р. Глубокая Щель, ущелье реки Западный Дагомыс), в типичных – 3 (ущелье р. Сочи, ущелье Ахцу на реке Мзымта и ущелье Шахгинское на реке Псоу) (рис. 1, табл. 1). Дискретность измерения метеорологических параметров составляла 3 часа и соответствовала срокам наблюдения на сетевых метеорологических станциях. Статистическая обработка полученных метеоданных проведена после экспорта результатов, накопленных регистраторами DS1923-F5, в MS Excel через программу OneWireViewer (НТЛ
«ЭлИн», Rus.).
Для сравнительной климатической характеристики исследуемой территории использовались данные гидрометеорологических станций первой категории: Анапа, Туапсе и Сочи, ряды наблюдений которых насчитывают до 130 значений за период с 1885 года, с небольшими перерывами во время первой и второй мировых войн. Кроме того, использованы данные станций: Новороссийск, Адлер (как самая южная точка). Для определения тенденций
- регионе использовались климатические нормы, установленные ВМО (осредненные характеристики за период 1960–1990 годов) (http://meteo.infospace.ru/win/). По значениям температуры воздуха (средние, максимальные, минимальные) и количеству выпавших
7
Туниев Б. С., Рыбак Е. А., Тимухин И. Н., Алиев Х. У.
Таблица 1
Районы изучения микроклимата на территории Сочинского национального парка
| Местоположение | Биохорион | Координаты WGS84 | Высота | ||
| № | н. у. м., | ||||
| (локалитеты) | Фитоландшафт | широта | долгота | ||
| м | |||||
| Окрестности | Восточно-Средиземноморский | ||||
| Пушистодубово- | |||||
| 1 | поселка | 44,014361 | 39,191878 | 96 | |
| можжевеловый сугрудок, | |||||
| Вишнёвка | |||||
| субтропический пинарий | |||||
| 2 | Ущелье реки | Восточно-Средиземноморский | 43,984583 | 39,2432 | 58 |
| Неожиданная | Широколиств. лес ущелья | ||||
| Окрестности | Восточно-Средиземноморский | ||||
| 3 | поселка | 43,877361 | 39,372486 | 33 | |
| Субтропический пинарий | |||||
| Солоники | |||||
| 4 | Гора Хакукай | Восточно-Средиземноморский | 44,024283 | 39,305131 | 599 |
| Можжевеловое редколесье | |||||
| Восточно-Средиземноморский | |||||
| 5 | Орлиные скалы | Скальный субтропический | 43,558842 | 39,821489 | 358 |
| пинарий | |||||
| 6 | Гора Большой | Восточно-Средиземноморский | 43,545469 | 39,850922 | 591 |
| Ахун | Субтропический пинарий | ||||
| 7 | Ущелье реки | Колхидский | 43,817017 | 39,447281 | 29 |
| Глубокая Щель | Лапинник | ||||
| Ущелье реки | Колхидский | ||||
| 8 | Западный | Полидоминантный | 43,763167 | 39,702353 | 336 |
| Дагомыс | колхидский лес | ||||
| Ущелье реки | Колхидский | ||||
| 9 | Полидоминантный | 43,713883 | 39,774869 | 234 | |
| Сочи | |||||
| колхидский лес | |||||
| Ущелье Ахцу | Колхидский | ||||
| 10 | Скально-лесной комплекс | 43,601292 | 40,02825 | 462 | |
| (р. Мзымта) | |||||
| известняков Колхиды | |||||
| Ущелье | Колхидский | ||||
| 11 | Шахгинское | Скально-лесной комплекс | 43,531814 | 40,095003 | 190 |
| (р. Псоу) | известняков Колхиды | ||||
атмосферных осадков были уточнены границы различных типов климата, а также климатических зон в пределах одного типа. Кроме того, определены современные тенденции изменения климатических характеристик в регионе исследования в XXI веке.
РЕЗУЛЬТАТЫ
Ранее были выполнены более масштабные климатические исследования на территории от Анапы до села Гячрыпш (быв. Леселидзе, Республика Абхазия) для определения условий произрастания сосны пицундской на Черноморском побережье (Джангиров, Шевцов, 2014). Климат исследуемого региона изменяется от умеренно-континентального, степного (Анапа), до влажного субтропического (Гячрыпш).
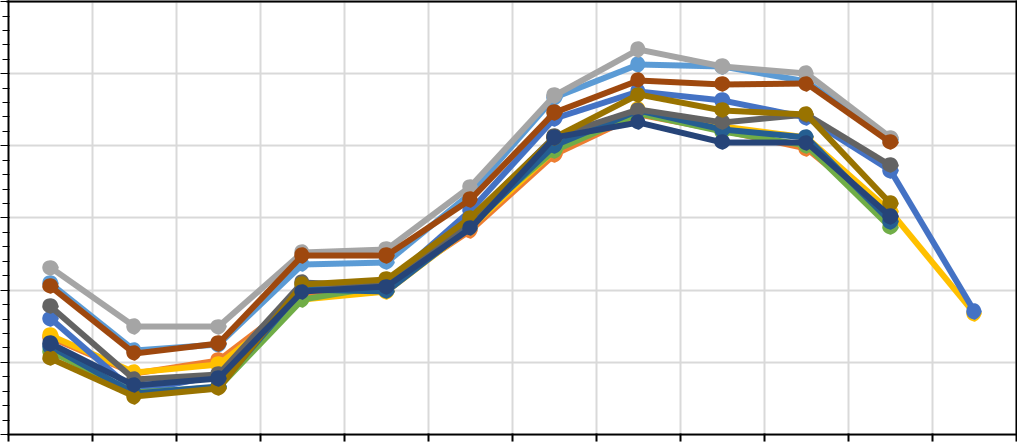
Большая часть территории российского побережья Кавкза характеризуется типично средиземноморским климатом (табл. 2). Подавляющая часть территории СНП лежит в зоне влажных субтропиков (табл. 2). Однако выполненные микроклиматические исследования показали её неоднородность. Полученные результаты по температуре и относительной влажности воздуха на изученных объектах приведены на рисунках 2 (а, b) и 3 (a, b). Кроме среднемесячных значений были расчитаны среднедекадные.
8
Хорологические закономерности в экотоне Восточно-Средиземноморской и Колхидской биогеографических провинций на территории Сочинского национального парка
![]()
Таблица 2
Климатическая характеристика Черноморского побережья российского Кавказа
| Температура, оС | Зона сухих субтропиков | Зона влажных субтропиков | |||||
| средиземноморского типа | |||||||
| и осадки, мм | |||||||
| Анапа | Новороссийск | Геленджик | Туапсе* | Лазаревское | Сочи | Адлер | |
| Средняя | 16,4 | 16,9 | 16,4 | 17,6 | 18 | 18,2 | 18,2 |
| максимальная | |||||||
| Среднегодовая | 12,4** | 13,1 | 13,7 | 13,2 | 13,8 | 14,1 | 13,5 |
| (13,6)*** | (13,9) | (13,8) | |||||
| Средняя | 9 | 11,1 | 10 | 9,7 | 10,3 | 10,8 | 9,4 |
| минимальная | |||||||
| Минимальная | −23,9 | −20,2 | −17,2 | −19 | −15 | −13,4 | −15 |
| Максимальная | 38,2 | 41 | 37,6 | 41,1 | 39 | 39,4 | 39 |
| Осадки | 560 | 758 | 716 | 1371 | 1541 | 1571,1 | 1341 |
| (563) | (825) | (756) | (1436) | (1371) | |||
Примечание к таблице. * Район южнее Туапсе расположен в зоне влажных субтропиков, что сильно отличает этот регион от более северного участка побережья от Анапы до Туапсе, где господствует типичный полусухой средиземноморский климат. Об этом свидетельствуют, прежде всего данные по количеству выпавших осадков, а также по значениям минимальных (средних и абсолютных) температур. ** Для Анапы, Туапсе и Сочи использованы климатические нормы; для остальных пунктов
– осредненные данные из литературных и интернет-источников (http://meteo.ru/pogoda-i-klimat).
*** В скобках для сравнения даны динамические климатические нормы – период осреднения 1981– 2010 года (https://meteoinfo.ru/climatcities).
Ниже приведены геоботанические и герпетофаунистические описания участков в местах изучения микроклимата.
Приморские ценозы восточно-средиземноморского типа описаны в Макопсинском участковом лесничестве СНП в окрестностях поселка Вишнёвка и в нижнем створе ущелья реки Неожиданная, а также в Лазаревском участковом лесничестве СНП – в окрестностях поселка Солоники.
Участок 1. В окрестностях поселка Вишнёвка (рис. 4a) представлены естественные и искусственные субтропические пинарии с Pinus pityusa Steven и P. pallasiana D. Don [Pinetum fruticosum], пушистодубово-можжевеловые сугрудки [Querceto pub. – Juniperetum fruticosum]
- доминированием Juniperus deltoides R. P. Adams, субдоминантом выступает Quercus pubescens Willd., при единичном участии Pinus pityusa, Fraxinus excelsior L., Carpinus orientalis Mill. Во втором ярусе доминирует Cotinus coggygria Scop., субдоминант – Ligustrum vulgare L. Здесь же развиты вторичные шибляки в местах былого культивирования садов, с
Juniperus deltoides, Paliurus spina-christi Miller, Swida australis (C.A. Mey.) Pojark. ex Grossh.,
Cotinus coggygria, Carpinus orientalis, Crataegus pentagyna Waldst. & Kit. ex Willd.,
- microphylla C. Koch, Pyracantha coccinea M. Roem, Mespilus germanica L., Ligustrum vulgare.
Внеярусная растительность составлена Periploca graeca L., Clematis vitalba L. и редко встречающимся Vitis sylvestris Gmel.
| oestrifera Bieb. | , | Anacamptis pyramidalis (L.) Rich. | , | Psoralea bituminosa ssp. pontica (A. P. | ||||
| Khokhr.) | Zernov, | Dorycnium | herbaceum Vill., | Astragalus circassicus Grossh., | ||||
| Scabiosa bipinnata K. Koch, S. columbaria L. и другие. | ||||||||
Для герпетофауны участка характерно преобладание восточно-средиземноморских видов (Tuniyev, 1995), в том числе Hyla orientalis Bedriaga 1890, Testudo graeca nikolskii Chkhikvadze et Tuniyev 1986, Pseudopus apodus (Pallas, 1775), Darevskia pontica (Lants et Cyren, 1919), Platyceps
9
| Туниев Б. С., Рыбак Е. А., Тимухин И. Н., Алиев Х. У. | ||||||||||||
| 30,0 | ||||||||||||
| 25,0 | ||||||||||||
| оС | 20,0 | |||||||||||
| , | ||||||||||||
| Температура | 15,0 | |||||||||||
| 10,0 | ||||||||||||
| 5,0 | ||||||||||||
| 0,0 | ||||||||||||
| 12 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |
| Вишневка | Макопсе | Солоники | ||||||||||
| Глубокая Щель | Хакукай | Западный Дагомыс | ||||||||||
| Ореховка | Орлиные скалы | Ахун | ||||||||||
| Ахцу | Псоу | |||||||||||
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
a
| % | 110 | |||||||||||
| 100 | ||||||||||||
| 90 | ||||||||||||
| 80 | ||||||||||||
| 70 | ||||||||||||
| 60 | ||||||||||||
| 50 | ||||||||||||
| 40 | ||||||||||||
| 12 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |
| Вишневка | Макопсе | Солоники | ||||||||||
| Глубокая Щель | Хакукай | Западный Дагомыс | ||||||||||
| Ореховка | Орлиные скалы | Ахун | ||||||||||
| Ахцу | Псоу | |||||||||||

b
Рис. 2. Среднемесячная температура (a) и относительная влажность воздуха (b) в местах наблюдения за микроклиматом в СНП
10
| Хорологические закономерности в экотоне Восточно-Средиземноморской и Колхидской | ||||||||||||
| биогеографических провинций на территории Сочинского национального парка | ||||||||||||
| 45,0 | ||||||||||||
| 40,0 | ||||||||||||
| 35,0 | ||||||||||||
| С | 30,0 | |||||||||||
| о, | ||||||||||||
| Температура | 25,0 | |||||||||||
| 20,0 | ||||||||||||
| 15,0 | ||||||||||||
| 10,0 | ||||||||||||
| 5,0 | ||||||||||||
| 0,0 | ||||||||||||
| 12 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |
| Вишневка | Макопсе | Солоники | ||||||||||
| Глубокая Щель | Хакукай | Западный Дагомыс | ||||||||||
| Ореховка | Орлиные скалы | Ахун | ||||||||||
| Ахцу | Псоу | |||||||||||
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
| a | ||||||||||||
| 45,0 | ||||||||||||
| 40,0 | ||||||||||||
| 35,0 | ||||||||||||
| С | 30,0 | |||||||||||
| о, | ||||||||||||
| Температура | 25,0 | |||||||||||
| 20,0 | ||||||||||||
| 15,0 | ||||||||||||
| 10,0 | ||||||||||||
| 5,0 | ||||||||||||
| 0,0 | ||||||||||||
| 12 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |
| Вишневка | Макопсе | Солоники | ||||||||||
| Глубокая Щель | Хакукай | Западный Дагомыс | ||||||||||
| Ореховка | Орлиные скалы | Ахун | ||||||||||
| Ахцу | Псоу | |||||||||||
| b | ||||||||||||
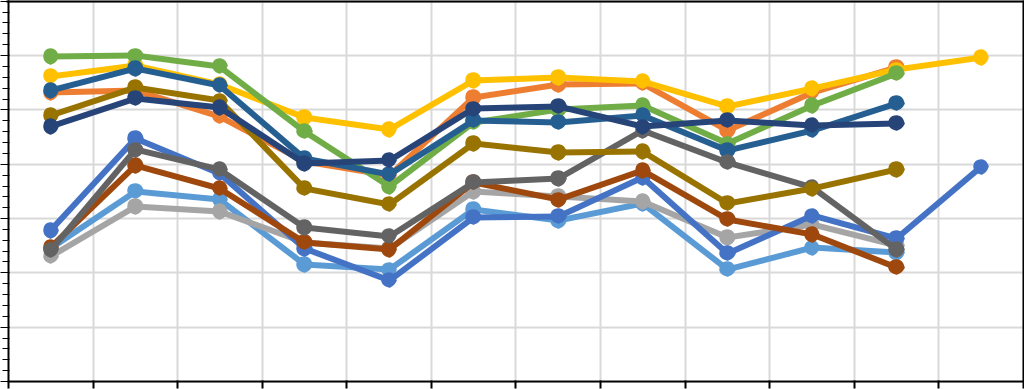
| Рис. 3. Амплитуда температуры (a) и относительной влажности воздуха (b) в местах | ||||||||||||
| наблюдения за микроклиматом СНП | ||||||||||||
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
11
Туниев Б. С., Рыбак Е. А., Тимухин И. Н., Алиев Х. У.
![]()
najadum (Eihwald, 1831), Dolichophis caspius (Gmelin, 1789). Из колхидских видов однократно отмечен Natrix megalocephala Orlov et Tuniyev, 1987.
Участок 2. Нижний створ ущелья реки Неожиданная (рис. 4b) занят грабинником грабовым тисово-плющовым [Carpineto – Carpinetum orien. taxoso-hederosum] на склоне юго-восточной экспозиции, крутизной 25о. Доминантом выступает Carpinus orientalis, субдоминант – Carpinus betulus L. Единично в древостое присутствуют Taxus baccata L., Fraxinus excelsior, Acer campestre L., A. cappadocicum Gled., Ulmus glabra Hudson, Sorbus torminalis (L.) Crantz, Cerasus avium (L.) Moench. Подлесок рассеянный, преимущественно образованный Ligustrum vulgare, Ilex colchica Pojark., при участии Crataegus microphylla, Staphylea colchica Steven, Swida australis, Euonymus leiophloea Steven, E. latifolius (L.) Mill. Травяно-кустарничковый ярус покрывает 85 %. В напочвенном покрове доминантом выступает Hedera helix L. – 60 %, отмечены Trachystemon orientalis (L.) G.Don. fil., Ruscus aculeatus L., Helleborus caucasicus A. Braun, Sanicula europaea L., Rubus caucasicus Focke, Potentilla micrantha Ramond ex DC., Brachypodium sylvaticum (Huds.) Beauv., Tamus communis L., Campanula rapunculoides L., Serratula quinquefolia M. Bied. ex Willd., Galebdolon luteum (L.) L., Aegonychon purpureocaeruleum (L.) Holub, Lathyrus laxiflorus (Desf.) O. Kuntze.
-
- ущелье имеется скальный участок с типичными представителями средиземноморской флоры: Colutea cilicica Boiss. et Balansa, Dianthus acantholimonoides Schischk., Teucrium chamaedrys L., Melampyrum arvense L., Seseli ponticum Lipsky, Aegonychon purpureocaeruleum
- другими, но основу растительности участка представляют широколиственные леса, развитие которых определяется узким ущельем и константным водотоком.
Герпетофауна участка представлена восточно-средиземноморскими (Hyla orientalis, Platyceps najadum, Dolichophis caspius), европейскими (Natrix natrix L., 1758), кавказскими (Rana macrocnemis Boul. 1885), и колхидскими (Darevskia brauneri (Méhelÿ, 1909)) видами.
Участок 3. Приморский склон в окрестностях поселка Солоники (рис. 4 c) занят одной из самых крупных старовозрастных рощ Pinus pityusa. Небольшие группы P. pityusa имеются и выше поселка Солоники, до 7 км уходящие вглубь по ориентированным на восток гребням хребтов правобережья реки Цусхвадж. В древостое практически по всей площади абсолютным доминантом, чаще монодоминантом, выступает Pinus pityusa. В верхней части сосняка местами субдоминантом выступает Quercus petraea L. ex Liebl., крайне редко, единичными деревьями, встречаются Cerasus avium, Carpinus betulus.
Рассматриваемый сосняк неоднороден, в зависимости от мощности почвенного слоя, выхода известняков на дневную поверхность, особенностей микрорельефа формируются парцеллы сосняка мертвопокровного [Pinetum nudum] и различных вариантов сосняка кустарникового [Pinetum fruticosum], с попеременным доминированием на более влажных участках и в балках – Staphylea colchica, Corylus avellana L., а на сухих участках – Rhus coriaria L., Cotinus coggygria. Все остальные древесно-кустарниковые породы либо встречаются рассеяно, либо формируют группировки на незначительных площадях, в том числе Ostrya carpinifolia Scop., Carpinus orientalis, Ligustrum vulgare, Crataegus microphylla, Paliurus spina-christi, Swida australis, Sorbus torminalis, Rubus caesius L., R. ibericus Juz., R. anatolicus (Focke) Focke ex Hausskn., Frangula alnus Mill., Rosa canina L., Chamaecytisus hirsutus (L.) Link. Внеярусная растительность развита в понижениях рельефа и представлена Hedera helix, Clematis vitalba, Smilax excelsa L., Periploca graeca, Vitis sylvestris, Lonicera caprifolium L.
-
- кустраничково-травяном ярусе доминируют Ruscus aculeatus, Brachypodium rupestre (Host) Roem. et Schult., субдоминантом в нижней части сосняка выступает
Dorycnium herbaceum, в верхней части – Pteridium tauricum V.I. Krecz. В травяном ярусе отмечены восточно-средиземноморские гемиксерофитные виды: Linum corymbulosum Reichenb., L. bienne Miller, L. tenuifolium L., Coronilla orientalis Miller, Sesleria alba Sm., Briza elatior Sibth. et Smith, Polygala major Jacquin, Jurinea arachnoidea Bunge, Muscari
armeniacum Leichtlin ex Baker, Poterium polygamum Waldst. & Kit., Orchis punctulata, Anacamptis pyramidalis, Ophrys oestrifera, Psoralea bituminosa ssp. pontica, Dictamnus albus L., Stachys recta L. subsp. atherocalyx (C.Koch) Derviz-Sokolova, Peucedanum tauricum M. Bieb.,
12
Хорологические закономерности в экотоне Восточно-Средиземноморской и Колхидской биогеографических провинций на территории Сочинского национального парка

Рис. 4. Ценозы восточно-средиземноморского типа на территории
Сочинского национального парка
Приморские ценозы: a – пушистодубово-можжевеловый сугрудок в окрестностях поселка Вишнёвка; b – грабинниково-грабовый лес в нижнем створе реки Неожиданная; c – сосняк коротконожковый в окрестностях поселка Солоники; Удаленные от приморской полосы локальные ценозы: d – можжевеловое редколесье на горе Хакукай; e – cосняк скальный в урочище Орлиные скалы; f – cосняк кустарниковый на горе Большой Ахун.
Seseli ponticum, Hieracium cymosum L., Scorzonera stricta Hornem., Blackstonia perfoliata (L.) Huds., Laser trilobum (L.) Borkh., Silene italica (L.) Pers., Cirsium euxinum Kharadze, Tamus communis, Dorycnium graecum (L.) Ser., Anthemis cotula L., Helianthemum nummularium (L.) Mill., Heracleum scabrum Albov, Teucrium chamaedrys, Aegonychon purpureocaeruleum, Erianthus ravennae (L.) P. Beauv., Convolvulus cantabrica L., Linaria genistifolia (L.) Mill., Verbascum gnaphalodes M. Bieb. и другие.
13
Туниев Б. С., Рыбак Е. А., Тимухин И. Н., Алиев Х. У.
![]()
Герпетофауна участка представлена восточно-средиземноморскими видами Pseudopus apodus, Darevskia pontica, Platyceps najadum, Dolichophis caspius, Natrix tessellata Laurenti, 1768. Колхидские (Ommatotriton ophryticus (Berthold, 1846) и кавказские (Rana macrocnemis Boulenger, 1885) виды присутствуют на участке в небольшой балке с ручьем, расположенной в центре лесного массива.
Удаленные от приморской полосы локальные восточно-средиземноморские ценозы описаны в Лыготхском участковом лесничестве СНП на горе Хакукай, а также в Мацестинском участковом лесничестве СНП – в урочище Орлиные скалы и на горе Большой Ахун.
Участок 4. На крутосклонных скальных обнажениях восточной экспозиции горы Хакукай (рис. 4d) сохранилось можжевеловое редколесье [Juniperetum saxatile] из Juniperus deltoides в окружении широколиственных, преимущественно дубовых лесов. Среди можжевельников встречается Carpinus orientalis, Swida koenigii (C.K. Schneid.) Pojark. ex Grossh., Cotinus coggygria, Ligustrum vulgare, Amelanchier ovalis Medik., Paliurus spina-christi, Jasminum fruticans L. Разреженный кустарничково-травяной покров составлен Ruscus aculeatus, Salvia ringens Sm., Teucrium chamaedrys, Laser trilobum, Peucedanum tauricum, Alyssum murale Waldst. & Kit., Helianthemum grandiflorum (Scop.) DC., Dianthus acantholimonoides, Asphodeline lutea (L.) Reichb.), Thymus helendzhicus Klokov & Des.-Shost., Sedum maximum (L.) Hoffm. subsp. caucasicum Grossh., Psephellus barbeyi Albov, Ceterach officinarum Willd., Asplenium ruta-muraria L., Anemone blanda Schott & Kotschy, Seseli ponticum, Scabiosa olgae Albov, Asperula abchasica V.I. Krecz., Aegonychon purpureocaeruleum, Carex cuspidata Host. и другие.
Для герпетофауны участка также характерно преобладание восточно-средиземноморских видов, в том числе Hyla orientalis, Testudo graeca nikolskii, Pseudopus apodus, Darevskia pontica, Platyceps najadum, Dolichophis caspius. Из колхидских и европейских видов, за пределами можжевелового редколесья, отмечены Anguis colchica (Nordmann, 1840), Natrix megalocephala, Zamenis longissimus Laurenti, 1768, Pelias kaznakovi (Nikolsky, 1909).
Участок № 5. В урочище Орлиные скалы (рис. 4e) сохранился сосняк скальный [Pinetum saxatile] естественного происхождения на скалах восточной экспозиции. В I ярусе доминирует Pinus pityusa, единично – Quercus petraea. Во II ярусе разреженно доминирует Cotinus coggygria, субдоминантом выступает Carpinus orientalis. Единично на участке отмечен в подросте Q. petraea, кустарники – Paliurus spina-christi, Rosa canina, Ligustrum vulgare, Crataegus microphylla, Juniperus deltoides, Cistus creticus L., Jasminum fruticans, Ficus carica L., имеющий здесь кустовую форму. В кустарничково-травяном покрове преобладает Brachypodium rupestre, реже встречается Peucedanum tauricum, Ruscus aculeatus, Dictamnus albus, Dorycnium graecum, D. herbaceum, Dianthus imereticus (Rupr.) Schischkin, Psoralea bituminosa L. subs. pontica, Psephellus barbeyi, Inula ensifolia L., I. salicina L. subsp. aspera (Poiret) Jav., Polygala major, Laser trilobum, Teucrium chamaedrys, Teucrium polium L., Convolvulus cantabrica, Sedum gracile C.A. Meyer, Asplenium ruta-muraria, Asphodelina lutea, Briza elatior, Euphorbia myrsinites L. и другие.
Герпетофауна участка представлена восточно-средиземноморскими видами – Hyla orientalis, Pseudopus apodus, Darevskia pontica, Platyceps najadum, Dolichophis caspius. За пределами участка, в лесных мезофильных биотопах обитают представители колхидских и европейских видов: Bufo verrucosissimus (Pallas, 1814), Rana macrocnemis, Darevskia brauneri, Coronella austriaca Laurenti, 1768, Zamenis longissimus, Pelias kaznakovi.
Участок № 6. В верхней трети горы Большой Ахун (рис. 4f) сохранился сосняк кустарниковый [Pinetum fruticosum] естественного происхождения с подсадкой лесных культур на склоне южной экспозиции, крутизной 10о. Древостой формирует Pinus pityusa, единично встречается Quercus iberica Steven. В подлеске содоминируют Cotinus coggygria, Crataegus microphylla, Swida australis, Carpinus orientalis, при участии Ligusticum vulgare, Cistus creticus, Rubus caucasicus, R. anatolicus, и в подросте – Sorbus torminalis, Tilia begoniifolia Stev. и Ulmus glabra. Внеярусная растительность представлена Smilax excelsa, Clematis vitalba, Periploca graeca, Lonicera caprifolium. В напочвенном травяном покрове
14
Хорологические закономерности в экотоне Восточно-Средиземноморской и Колхидской биогеографических провинций на территории Сочинского национального парка
![]()
доминирует Epimedium pinnatum subsp. colchicum, субдоминантом выступает Trachystemon orientalis. Также встречаются Ruscus aculeatus, Pteridium tauricum, Platantera chloranta (Cust.) Reichend., Cephalanthera damasomium (Mill.) Druce, Brachypodium rupestre, Dictamnus albus, Iris colchica Kem.-Nath., Tamus communis, Euphorbia amygdaloides L., E. squamosa Willd., Thalictrum minus L., Lathyrus laxiflorus, L. vernus (L.) Bernh., Colchicum umbrosum Steven, Viola dehnhardtii Ten., Carex cuspidata, Aristolochia steupii Woronow, Psoralea bituminosa ssp. pontica, Galega officinalis L., Veronica umbrosa M. Bieb. и другие.
Герпетофауна участка представлена средиземноморскими (Hyla orientalis, Pseudopus apodus, Platyceps najadum, до конца ХХ века на участке единично попадалась Testudo graeca nikolskii) и европейскими (Anguis fragilis, Coronella austriaca) видами. За пределами участка встречаются колхидские виды – Darevskia derjugini (Nikolsky, 1898), Zamenis longissimus.
Колхидские ценозы на западной границе хориона описаны в Головинском участковом лесничестве СНП в нижней части ущелья реки Глубокая Щель и в Дагомысском участковом лесничестве СНП в ущелье реки Западный Дагомыс.
Участок № 7. В нижней части ущелья реки Глубокая Щель (рис. 5a) развит лапинник разнотравный [Pterocarietum variaherbosum]. Сомкнутость древесного яруса 95 %. Доминантом выступает Pterocarya fraxinifolia (Lam.) Spach, единично присутствуют Acer campestre, Alnus glutinosa (L.) Gaertin., Ficus carica. В подросте отмечены Pterocarya fraxinifolia, Fraxinus exselsior, Carpinus betulus, Acer campestre, A. pseudoplatanus L., Ficus carica, Dyospiros lotus L. Подлесок составлен Corylus avellana, Swida australis, Sambucus nigra, Euonymus europaeus L., Staphylea colchica, Crataegus microphylla. Внеярусная растительность представлена Hedera helix, Smilax excelsa.
Кустарничково-травяной ярус занимает 50 % площади, явные доминанты отсутствуют. Отмечено произрастание Rubus caucasicus, Rubus caesius, Oplismenus undulatifolius (Ard.) P. Beauv., Galeobdolon luteum, Pimpinella sp., Symphytum grandiflorum, Phyllitis scolopendrium (L.) Newman, Polystichum setiferum (Forssk.) Moore ex Woyn., Dryopteris filix-mas (L.) Schott, Trachystemon orientalis, Carex pendula Huds., Helleborus caucasicus, Sanicula europaea, Circaea lutetiana L., Potentilla micrantha, Primula vulgaris, Carpesium cernuum L., Tamus communis, Arum orientale, Aristolochia steupii, Sium sisarum L. Близость автомобильных дорог и поселка Глубокая Щель определяют наличие сорных и чужеродных видов – Geum urbanum L., Urtica dioica L., Duchesnea indica (Andrews) Focke, Bidens tripartita L.
Герпетофауна участка представлена колхидскими и кавказскими видами – Bufo verrucosissimus, Rana macrocnemis, Darevskia brauneri, Natrix megalocephala, Zamenis longissimus, при наличии европейских и европейско-средиземноморских видов – Hyla orientalis, Pelophylax ridibundus (Pallas, 1771), Anguis colchica.
Участок 8. В узком ущелье среднего течения реки Западный Дагомыс (рис. 5b) развит полидоминантный колхидский лес с вечнозеленым подлеском. Микроклимат записывался на склоне восточной экспозиции, на участке грабо-букняка самшитового [Fagetum carpinoso-buxosum]. Доминантом в I ярусе является Fagus orientalis, субдоминант – Carpinus betulus, при участии Tilia begoniifolia, Fraxinus excelsior, Acer pseudoplatanus, A. campestre, Alnus barbata. Во втором ярусе доминируют Buxus colchica, присутствуют Taxus baccata, Ostrya carpinifolia, Ficus carica, Acer laetum C.A. Mey., в кустарниковм ярусе – Swida australis, Daphne pontica L., Ilex colchica, Euonymus latifolia, Philadelphus caucasicus Koehne, Laurocerasus oficinalis M. Roem., Staphylea colchica, Sambucus nigra, Rhododendron ponticum L., Rh. luteum. Внеярусная растительность составлена Periploca graeca, Hedera colchica, Clematis vitalba, Vitis sylvestris, Lonicera caprifolium.
-
- кустарничково-травяном ярусе представлены Ruscus colchicus, Epimedium pinnatum subsp. colchicum, Omphalodes сappadocia (Willd.) DC., Lilium martagon L. subsp. caucasicum Miscz. ex Grossh., Paeonia caucasica (Schipcz.) Schipcz., P. wittmanniana Hartwiss ex Lindl., Galanthus woronowii, Cyclamen coum, Ophioglossum vulgatum L., Asplenium adiantum-nigrum L., A. trichomanes, Phyllitis scolopendrium, Pteris cretica L., Adiantum capillus-veneris L., Orchis mascula, Dactylorhiza urvilleana, Helleborus caucasicus, Scilla bifolia, Vinca pubescens D’ Urville, V. minor L., Aristolochia steupii, Arum orientale, Alliaria petiolata (M. Bieb.) Cavara & Grande,
15
Туниев Б. С., Рыбак Е. А., Тимухин И. Н., Алиев Х. У.

Рис. 5. Колхидские ценозы на территории Сочинского национального парка
Ценозы на западной границе хориона: a – лапинник в нижнем створе ущелья Глубокая Щель; b – широколиственный лес с вечнозеленым подлеском в ущелье реки Западный Дагомыс; Типичные колхидские ценозы центральной части Колхидского биохориона: c – ущелье реки Сочи; d – ущелье Ахцу на реке Мзымта; e, f – Шахгинское ущелье на реке Псоу.
Colchicum umbrosum, C. speciosum, Paris incompletа M. Bieb., Arabis nordmanniana Rupr., Polygonatum orientale Desf., P. glaberrimum K. Koch, Corydalis caucasica, Umbilicus oppositifolius Ledeb., Scopolia carniolica Jacq., Atropa caucasica Kreyer и другие.
- составе герпетофауны преобладают колхидские виды – Ommatotriton ophryticus, Bufo berrucosissimus, Rana macrocnemis, Pelodytes caucasicus Boul., 1896, Darevskia brauneri, Natrix megalocephala, Zamenis longissimus, Pelias kaznakovi, присутствуют европейские и
16
Хорологические закономерности в экотоне Восточно-Средиземноморской и Колхидской биогеографических провинций на территории Сочинского национального парка
![]()
европейско-средиземноморские виды – Hyla orientalis, Pelophylax ridibundus, Anguis colchica, Coronella austriaca, Natrix tessellata.
Типичные колхидские ценозы центральной части Колхидского биохориона описаны в Верхне-Сочинском участковом лесничестве в ущелье реки Сочи в окрестностях села Ореховка, в Кепшинском участковом лесничестве – в ущелье Ахцу на реке Мзымта и в Весёловском участковом лесничестве в ущелье Шахгинское на реке Псоу.
Участок 9. Ущелье реки Сочи в окрестностях села Ореховка покрыто богатыми по составу широколиственными лесами колхидского типа с вечнозеленым подлеском (рис. 5c). На отдельных участках можно выделить дубняк кустарниково-зверобойный [Quercetum hypericosum xylost.], буко-каштанник понтийско-рододендроновый [Fageto-Castanetum rododendrosum pont.], грабо-букняк самшитовый [Carpineto-Fagetum buxosum]. Микроклимат участка изучен по правому борту ущелья, на склоне 25° восточной и юго-восточной экспозиций. В древесном ярусе, в зависимости от локальных условий, доминантами выступают Fagus orientalis, Quercus iberica, Carpinus betulus, Castanea sativa. В I ярусе присутствуют Fraxinus excelsior, Tilia begnoniifolia, Diospyros lotus, Alnus barbata, Acer pseudoplatanus, A. campestre, A. laetum, Cerasus avium. В кустарниковом ярусе в дубняке доминирует Hypericum xylosteifolium (Spach) N. Robson, субдоминантом является Rhododendron luteum; в букняках – попеременно Rhododendron ponticum и Buxus colchica, при значительном участии Laurocerasus officinalis, Ilex colchica, Staphylea colchica, Corylus avellana, Sambucus nigra, Ruscus colchicus и спорадичной встречаемости Euonymus latifolia, E. leiophloea, E. europaeus, Daphne pontica, Hypericum androsaemum L., Leptopus colchicus (Fischer et C.A.Meyer ex Boiss.) Pojark. (западная граница ареала вида).
- травяном покрове отмечены Ophioglossum vulgatum, Pteris cretica, Blechnum spicant (L.), Adiantum capillus-veneris, Asplenium adiantum-nigrum, A. trichomanes, Phyllitis scolopendrium, Woodsia fragilis (Trev.) Moore, Epimedium pinnatum subsp. colchicum, Omphalodes сappadocia, Paeonia caucasica, Galanthus woronowii, Cyclamen coum, Orchis mascula, Helleborus caucasicus, Scilla bifolia, Vinca pubescens, V. minor, Aristolochia steupii, Symphitum grandiflorum, Arum orientale, Colchicum umbrosum, Carex pendula, Paris incompletа, Potentilla micrantha, Tamus communis, Arabis nordmanniana, Corydalis caucasica, Umbilicus oppositifolius, Scopolia carniolica, Euphorbia amygdaloides, Trachystemon orientalis, Lathyrus vernus и другие.
- составе герпетофауны преобладают колхидские виды – Ommatotriton ophryticus, Bufo berrucosissimus, Rana macrocnemis, Pelodytes caucasicus, Darevskia brauneri, D. derjugini, Natrix megalocephala, Zamenis longissimus, присутствуют европейские и европейско-средиземноморские виды – Hyla orientalis, Pelophylax ridibundus, Anguis colchica, Coronella austriaca, Natrix tessellata.
Участок 10. Ущелье Ахцу – уникальный рефугиум скально-лесной флоры и растительности известняковой Колхиды, являющийся частью Абхазского флористического района (Тимухин, Туниев, 2016) (рис. 5d). Богатая флора включает типичные колхидские и общекавказские виды с достаточно высоким представительством высокогорных видов
(Asplenium woronowii H. Christ, Cystopteris fragilis (L.) Bernh., Poa alpina L., Anthemis triumfetti (L.) All., Cirsium ciliatum (Murray) Moench,, Psephellus hypoleucus (DC) Boiss., Onosma caucasica Levin ex M.Pop., Campanula dzyschrica Kolak., Lonicera steveniana, Cerastium ponticum Albov и другие) и узких северо-колхидских (абхазских) эндемов (Campanula sclerophylla (Kolak.) Oganesian, Bupleurum rischawii Albov, Seseli rupicola Woronow, Kemulariella abchasica (Kem.-Nath.) Tamamsch., Muscari dolychanthum Woronow et Tron., Leptopus colchicus, Genista kolakowskyi Sachok., Dioscorea caucasica Lipsky и другие) (Тимухин, Туниев, 2018).
Древесные виды скальной растительности представлены аборигенными Fraxinus excelsior, Acer campestre, Acer sosnowskyi Doluch., Carpinus betulus, С. orientalis, Castanea sativa, Fagus orientalis, Quercus petraea, Cerasus avium, Populus alba L., Ostrya carpinifolia, Diospyros lotus, Taxus baccata, Tilia begoniifolia и чужеземными – Morus alba L., Paulownia tomentosa (Thun.) Steud. и другими; кустарники – Corylus avellana, Buxus colchicus, Euonymus leiophloea, Cornus mas, Swida koengii, S. australis, Juniperus communis L. ssp. oblonga (Bieb.) Galushko, Leptopus colchicus, Philadelphus caucasicus, Ligusticum vulgare, Cotoneaster
17
Туниев Б. С., Рыбак Е. А., Тимухин И. Н., Алиев Х. У.
![]()
integerrimus Medicus, Pyracantha coccinea, Staphylea colchica, Daphne pontica и другими. Внеярусная растительность составлена характерными для региона лианами: Periploca graeca, Hedera colchica, H. helix, Clematis vitalba, Smilax exelsa, Lonicera caprifolium. Особенности травяного покрова охарактеризованы выше.
Герпетофауна участка представлена типичными колхидскими и кавказскими видами Darevskia brauneri, Zamenis longissimus, Natrix megalocephala, Pelias kaznakovi, а также европейскими видами – Anguis colchica, Coronella austriaca. Из средиземноморских видов в нижней части ущелья отмечен Natrix tessellata.
Участок 11. Шахгинское ущелье реки Псоу – это наиболее представительный в СНП и России участок колхидской флоры и растительности, расположенный на границе с Республикой Абхазия (рис. 5e). Восточные склоны хребтов Дзыхра и Ахцу формируют правый (российский) борт ущелья
- лесными и скально-лесными комплексами, идентичными по составу с ущельем Ахцу, но с менее развитыми открытыми скальными ландшафтами. По днищу ущелья развит полидоминантный лес с вечнозеленым подлеском: буко-каштанник самшитовый [Fageto-Castanetum buxosum], грабо-букняк самшитовый [Carpineto-Fagetum buxosum]. В древесном ярусе, в зависимости от локальных условий, доминантами выступают Fagus orientalis, Carpinus betulus, Castanea sativa. В I ярусе присутствуют Fraxinus excelsior, Tilia begnoniifolia, Alnus barbata, Acer pseudoplatanus, A. campestre, A. laetum, Cerasus avium, Ostrya carpinifolia. В подлеске доминирует Buxus colchica, при значительном участии Laurocerasus officinalis, Ilex colchica, Staphylea colchica, Corylus avellana, Sambucus nigra, Ruscus colchicus и спорадичной встречаемости Euonymus latifolia,
E. leiophloea, E. europaeus, Daphne pontica, Hypericum androsaemum, Ruscus colhicus.
- травяном ярусе – Cyclamen coum, Helleborus caucasicus, Lusula forsteri, Limodorum
abortivum, Platanthera chlorantha, Listera ovata, Orchis mascula, Colchicum umbrosum, С. speciosum, Oplismenus undulatifolius, Saxifraga cymbalaria L., Aristolochia steupii, A. iberica и другие.
- сложеннии скальной флоры принимают участие Acer sosnowskyi, Taxus baccata, Ostrya carpinifolia, Buxus colchica, Staphylea colhica, Ficus carica, Swida koenigii, Leptopus colchicus, Genista kolakowskyi, Tamus communis, Seseli petraeum, Skabiosa olgae, Blackstonia perfoliata, Gentiana paradoxa, Mycelis muralis, Asplenium adiantum-nigrum, A. ruta-muraria, A. trichomanes, Ceterach officinarum, Pteris cretica, Campanula alliariifolia, C. pendula, Arabis caucasica, Ranunculus suukensis и другие. По всему ущелью отмечены реликтовые участки произрастания Dioscorea caucasica, характерна частая встречаемость Vitis sylvestris. Это единственное на территории РФ место произрастания Potentilla kamilae Kolak. и второе, после ущелья Ахцу, место произрастания в РФ Bupleurum rischawii.
Герпетофауна участка представлена типичными колхидскими и кавказскими видами
Ommatotriton ophryticus, Bufo berrucosissimus, Rana macrocnemis, Pelodytes caucasicus, Darevskia brauneri, D. derjugini, Natrix megalocephala, Zamenis longissimus, Pelias kaznakovi,
присутствуют европейские и европейско-средиземноморские виды – Hyla orientalis, Pelophylax ridibundus, Anguis colchica, Coronella austriaca.
ОБСУЖДЕНИЕ
- наличии средиземноморских видов в Западном Закавказье известно давно, начиная с классической работы Н. И. Кузнецова (1891). Описанные нами в СНП восточно-средиземноморские фитоландшафты и фауна населяющих их представителей амфибий и рептилий отличаются высоким сходством, независимо от географического расположения. Для большинства исследованных участков характерны такие представители древесно-кустарниковых видов, как Pinus pitysa, Juniperus deltoides, Ostrya carpinifolia, Paliurus spina-christi, Cotinus coggygria, Rhus coriaria, Carpinus orientalis, Pyracantha coccinea, Ligustrum vulgare, Jasminum fruticans. Во флоре кустарничково-травянистого яруса также характерны восточно-средиземноморские представители, практически отсутствующие в окружающих ценозах колхидского типа: Ruscus aculeatus, Asphodeline lutea, Orchis purpurea, O. tridentata, Ophrys oestrifera, Anacamptis pyramidalis, Psoralea bituminosa ssp. pontica, Dorycnium herbaceum, D. graecum, Scabiosa bipinnata, Linum
18
Хорологические закономерности в экотоне Восточно-Средиземноморской и Колхидской биогеографических провинций на территории Сочинского национального парка
![]()
corymbulosum, L. bienne, L. tenuifolium, Coronilla orientalis, Brachypodium rupestre, Briza elatior, Sesleria alba, Stachys recta subsp. atherocalyx, Peucedanum tauricum, Laser trilobum, Anthemis cotula, Helianthemum nummularium, Teucrium chamaedrys, Aegonychon purpureocaeruleum, Convolvulus cantabrica, Linaria genistifolia и другие. В составе герпетофауны этих участков также преобладают восточно-средиземноморские виды: Hyla orientalis, Testudo graeca nikolskii, Pseudopus apodus, Darevskia pontica, Platyceps najadum, Dolichophis caspius.
Некоторые различия в составе флоры восточно-средиземноморских видов наблюдаются
- западной и восточной частях СНП, одним из таких стыков биохорионов в СНП является гора Бозтепе на водоразделе рек Псезуапсе и Аше (Тимухин, 2008). Практически не выходят за пределы Восточно-Средиземноморской биогеографической провинции такие виды, как
Quercus pubescens, Colutea cilicica, Dianthus acantholimonoides, Astragalus circassicus, Thymus gelendzhicus, Salvia ringens, Scabiosa columbaria, Scorzonera stricta, Stypa pulcherima K.Koch,
Orchis punctulata, O. militaris, и сегодня определяющие границу этого биохориона.
- восточной части СНП в составе восточно-средиземноморских ценозов встречаются термофильные виды, отсутствующие в условиях воздействия холодных зимних воздушных масс в западной части СНП. К последним относятся Cistus creticus L., C. salviifolius L., Cytisus monspessulanus L., Euphorbia myrsinites и, встречавшийся до недавнего времени в СНП (Алиев и др., 2020), Arbutus andrachne L.
Как на востоке, так и на западе СНП к восточно-средиземноморским ценозам приурочены виды герпетофауны средиземноморского корня, часть которых может незначительно проникать в соседние колхидские ценозы, но колхидские виды амфибий и рептилий практически отсутствуют в гемиксерофильных биотопах. При этом, только в восточной части ареала, начиная с горы Большой Ахун и далее до Абхазии, встречается меланистическая форма Platyceps najadum (Tuniyev et al., 2019).
Представляет интерес определение факторов среды, способствующих сохранению восточно-средиземноморской биоты в целом и её спорадично вписанных в окружающие колхидские ценозы эксклавов в пределах Колхидской биогеографической провинции. Анализ полученных данных по микроклимату изученных 11 участков территории СНП показал, что при общих трендах изменения температуры и влажности воздуха для всей территории СНП, участки, корреспондирующие условиям восточно-средиземноморских ландшафтов, отличаются по ходу показателей температуры и влажности воздуха.
Основными климатообразующими факторами рассматриваемого региона являются солнечная
радиация, циркуляция атмосферы и характер подстилающей поверхности (Рыбак, 2009).
- формировании климата важную роль играет рельеф, под влиянием которого видоизменяется циркуляция воздушных масс. Особенно большое влияние на климат оказываeт Большой Кавказ, заслоняющий территорию в широтном направлении, препятствуя свободному переносу воздушных масс с севера на юг, и, до бассейна реки Псезуапсе, служит практически непреодолимым барьером для вторжений холодных воздушных масс с севера и северо-востока. Западнее, в бассейне реки Аше, хребет понижается и осенне-зимние холодные северо-восточные воздушные массы (бора) прорываются в среднем 4 дня в году. Уже этого непродолжительного периода холодного фронта становится достаточно для выпадения наиболее теплолюбивых колхидских видов растений и животных.
Открытость влажным южным и западным ветрам с Черного моря и защищенность района Сочи горными хребтами с трех сторон создает благоприятные условия для стационирования поступающих воздушных масс, главным образом, с запада, которые способствуют интенсивному формированию кучево-дождевых облаков, дающих обильные осадки и сохранению относительно высоких температур воздуха, характерных для подавляющей части территории СНП.
Представляется, что механизм влияния глобального потепления на количество осадков в масштабе относительно небольшой территории связан с изменениями в режиме атмосферной циркуляции. Исследуемый регион находится под влиянием Североатлантического колебания, обладающего собственной квазициклической изменчивостью (Рыбак, Рыбак, 2009). По всей видимости, в настоящее время изменчивость годовых сумм осадков в регионе Черного моря
19
Туниев Б. С., Рыбак Е. А., Тимухин И. Н., Алиев Х. У.
![]()
обусловлена собственной изменчивостью циркуляции атмосферы над Европой, на которую накладываются локальные факторы, и в гораздо меньшей степени связана с текущими изменениями глобальной температуры воздуха.
Было высказано не подкрепленное инструментально мнение (Скворцов, Рогова, 2020), что на высотное распространение термофильных видов в Колхиде влияет не только уменьшение температуры с подъемом в горы, но и другие факторы, позволяющие при наличии равновысоких участков, сохранение более теплого и мягкого климата на расположенных ближе к побережью участках. При одинаковых климатических параметрах локальные склоны, обращенные к югу, имеют более благоприятные температурные условия, благодаря которым термофильная растительность поднимается выше.
- целом, можно согласиться с этим очевидным фактом, однако наше исследование показало, что для средиземноморских видов большее значение имеет влажность воздуха. Анализ среднемесячных значений температуры и относительной влажности воздуха в 11 точках наблюдения за микроклиматом, несмотря на синхронный ход в течение года, отчетливо показал наличие в СНП двух совершенно различных микроклиматических групп (рис. 2 и 3). Особенно это проявляется в распределении относительной влажности воздуха. В таблице 3 представлены обобщенные характеристики этих двух микроклиматических групп.
Таблица 3
Среднемесячные показатели в двух микроклиматических группах
| Температура воздуха, оС | Относительная влажность воздуха, % | |||||||
| Группа | Средняя за | Средняя | Средняя | |||||
| период | Макс. | Мин. | амплитуда | Среднемесячная | Макс. | Мин. | амплитуда | |
| наблюдения | ||||||||
| 1 | 15,4 | 43,5 | -9,5 | 19,2 | 73,7 | 100 | 7,6 | 71,1 |
| 2 | 13,4 | 34,5 | -8,5 | 17,8 | 81,7 | 100 | 13,5 | 54,1 |
При этом, в границах Восточно-Средиземноморской биогеографической провинции, узкие лесные ущелья (например, р. Неожиданная) в границах зонального Средиземноморья имеют ход температуры и влажности воздуха, слабо отличающийся от таковых в Колхиде.
- изолированные реликтовые восточно-средиземноморские ценозы внутри Колхиды, сохраняющиеся в условиях эдафической сухости на скалах восточной экспозиции (г. Хакукай, Орлиные скалы, г. Большой Ахун), по ходу температуры и влажности воздуха практически не отличаются от приморских участков зонального Средиземноморья.
Кроме микроклиматических наблюдений в двух выявленных группах СНП, для реализации поставленной цели нами были проанализированы длительность периодов без осадков, а также выявлены периоды, когда количество выпавших атмосферных осадков было меньше климатической нормы (табл. 4). Отметим, что период наблюдений за микроклиматом (с декабря 2019 по ноябрь 2020 года) выдался очень засушливым.
Известно, что известняки обладают высокой термоёмкостью и фильтрующей влагу способностью. Эти особенности позволяют сохранение небольших по площади, но типичных по составу флоры и фауны участков восточно-средиземноморской биоты в окружении колхидской биоты. Аналогичные участки наблюдаются и восточнее, вплоть до территории Абхазии, где в центре известняковой Колхиды (бассейн р. Бзыбь) также наблюдается высокое представительство восточно-средиземноморских видов флоры и их сообщества, главным образом в скальных биотопах (Алиев и др., 2020). Современная сохранность этих реликтовых участков позволяют судить о былых размерах Черноморского ксерофильного рефугиума
(Tuniyev, 1995).
20
Хорологические закономерности в экотоне Восточно-Средиземноморской и Колхидской биогеографических провинций на территории Сочинского национального парка
![]()
Сопостовимые материалы получены в Западном Средиземноморье на примере
| финикийского | можжевельника | (Juniperus phoenicea L.) (Salvà-Catarineu, 2021), где его | |||
| Таблица 4 | |||||
| Длительность периодов без осадков и с осадками меньше климатической нормы | |||||
| Осадки, мм | Дни без осадков, шт | ||||
| Метеостанции | за период | за теплый | за период | за теплый период | |
| период | |||||
| наблюдений | наблюденийй | наблюдений | |||
| наблюдений | |||||
| Туапсе | 997 | 444 (275*) | 132 (37 %) | 80 (37 %) | |
| Сочи | 1120 | 364 (468*) | 135 (30 %) | 80 (37 %) | |
Примечание к таблице. * дефицит осадков (от среднего за 2000–2020 годы).
распространение на Пиренейском полуострове связано со средиземноморским типом биоклимата, ксеро-океаническим и полисезонно-океаническим подтипом, мезо- и надсредиземноморским термотипом и полузасушливыми, от сухих до субгумидных омбротипов (Rivas-Martínez et al., 2017). При этом, на юге Франции и севере Италии J. phoenicea встречается в климатических условиях, аналогичных средиземноморскомур егиону, но также растет в регионах с умеренным климатом на определенных участках, таких как крутые каменистые склоны гор, обращенные на юг (Mandin, 2005). Вид адаптирован к средиземноморскому климату и к широкому спектру биоклиматов, от субаридных до субгумидных или даже влажных, в мезо-средиземноморской, надсубсредиземноморской и оро-субсредиземноморской зонах (Mazur et al., 2016; Rivas-Martínez et al., 2004 г.).
Давая оценку возможным перспективам расширения ареалов средиземноморских видов
- существующем экотоне Средиземноморья и Колхиды отметим, что с 1970-х годов наблюдается монотонный рост глобальной и полушарной температур. Линейный тренд среднегодовой температуры за период 1976–2020 годов составил для Земного шара + 0,18 оС /
10 лет (объясненная трендом доля дисперсии ряда – 87 %), для Северного полушария: + 0,34 оС / 10 лет (86 %). Для России в целом среднегодовая аномалия температуры составила + 3,47 оС – максимальная величина в ранжированном по убыванию ряду наблюдений (предыдущий рекорд 2007: это на целый градус выше предыдущего максимума, зафиксированного в 2007 году) (Доклад об особенностях…, 2021). На уровне регионов обнаруживается серьезная пространственная и временная неоднородность в тенденциях изменений климата, при этом, как правило, с уменьшением масштаба пространственного осреднения заметно увеличивается как масштаб величины аномалий температуры воздуха, так и их межгодовая изменчивость (Груза, Ранькова, 2014). Кроме того, на фоне роста глобально-осредненной температуры может наблюдаться и ее снижение в отдельных регионах (Rybak, Rybak, 2017). Аналогичные тенденции выявлены и на всей территории СНП, а также по всему побережью Краснодарского края.
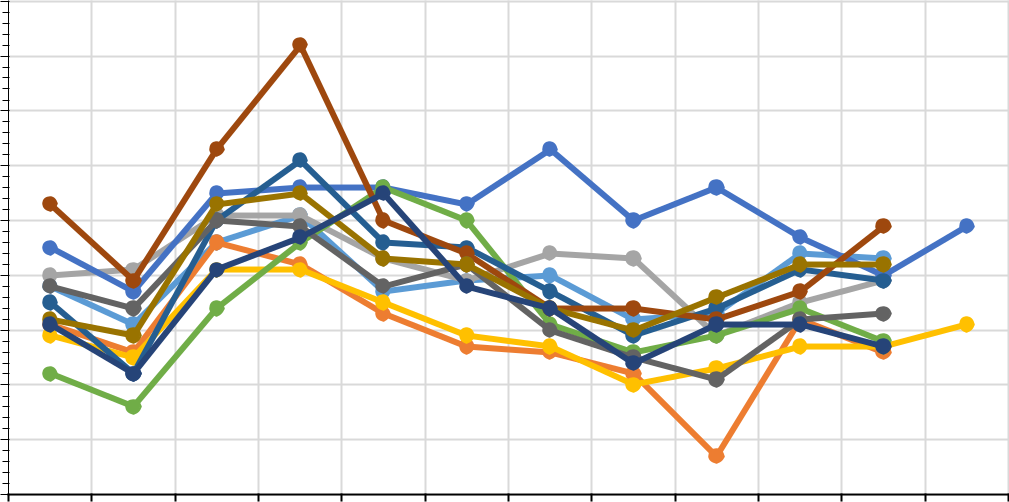
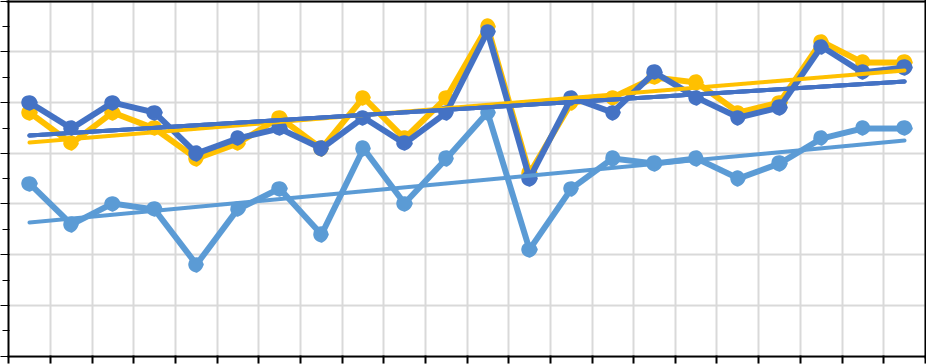
На рисунке 6 представлено распределение среднегодовой температуры воздуха (а) и годовая сумма атмосферных осадков (b) с 1999 по 2020 год на трех основных станциях.
На первый взгляд, в изменениях метеорологических характеристик на вышеуказанных станциях нет ничего экстраординарного. Выделенные тенденции хорошо согласуются с результатами других исследователей (Пестерева, 2011; Рыбак, Рыбак, 2013; Алешина и др., 2019 и др.), эксперты по всему миру признают факт глобального потепления. Однако, отметим две особенности: четко выделяются станции, расположенные в различных климатических зонах; среднегодовые температуры растут и растут практически синхронно. На фоне увеличения годовых сумм осадков на территории России в целом (Доклад об особенностях…, 2021), в Анапе их годовая сумма практически не изменяется; а в районе Туапсе и Сочи – отчетливая синхронная тенденция их уменьшения. Если в случае температуры воздуха анализ рядов позволяет выделить однозначный вывод об изменениях термического режима
21
| Туниев Б. С., Рыбак Е. А., Тимухин И. Н., Алиев Х. У. | |||||||||||
| (потепления) в регионе, то в случае с атмосферными осадками картина более сложна. | |||||||||||
| Обусловлено это, прежде всего, более сильной межгодовой изменчивостью годовых сумм | |||||||||||
| осадков на каждой отдельной метеостанции региона. | |||||||||||
| 17,0 | |||||||||||
| 16,0 | |||||||||||
| °С | 15,0 | ||||||||||
| Температура | 14,0 | ||||||||||
| 13,0 | |||||||||||
| 12,0 | |||||||||||
| 11,0 | |||||||||||
| 10,0 | |||||||||||
| 1999 | 2001 | 2003 | 2005 | 2007 | 2009 | 2011 | 2013 | 2015 | 2017 | 2019 | |
| Анапа | Туапсе | Сочи | |||||||||
| Линейная (Анапа) | Линейная (Туапсе) | Линейная (Сочи) | |||||||||
| a | |||||||||||
![]()
![]()
![]()
![]()
![]()
![]()
![]()
мм
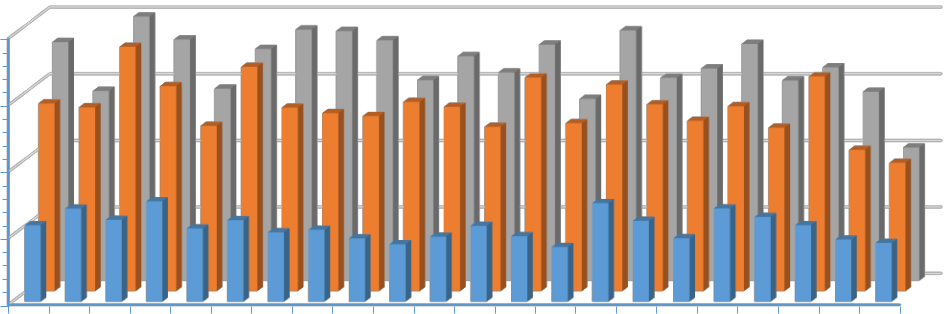
2000
1500
1000
500
0
| 1999 | 2000 | 2001 | 2002 | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 |
![]() Анапа Туапсе
Анапа Туапсе ![]() Сочи
Сочи
b
Рис. 6. Распределение среднегодовых температур воздуха (a) и годовых сумм
атмосферных осадков (b) за период 1999–2020 годов
На пространственную структуру поля осадков оказывают влияние помимо особенностей крупномасштабных синоптических процессов также и локальные факторы, например, рельеф местности (Rybak et al., 1994). Совокупное влияние локальных и нелокальных факторов приводит не только к систематическим изменениям в структуре поля осадков, но и к пространственным изменениям его вариаций.
Результаты прогностического моделирования (Егошин, 2021) указывают на повышение среднегодовой температуры в Сочинском регионе к 2050 году от 1,5 до 2,6 °C, а к 2070 году от 1,6 до 3,6 °С. При этом рост максимальной температуры самого тёплого месяца года к 2050 году составит от 2,3 °C до 3,6 °C, а к 2070 году составит 2 °C и 5 °C, соответственно, и годовая
22
Хорологические закономерности в экотоне Восточно-Средиземноморской и Колхидской биогеографических провинций на территории Сочинского национального парка
![]()
сумма осадков будет сокращаться, максимально от − 42 мм к 2050 году до − 36 мм к 2070 году.
-
- этими данными согласуются наши материалы по пространственному изменению ареалов и численности представителей колхидской и восточно-средиземноморской эколого-географических групп амфибий и рептилий. Так, Testudo graeca nikolskii отмечена в ряде районов на удалении от приморской полосы (на хребте Бачко, на водоразделе бассейнов рек Цусхважд и Чухукт, на склонах горы Жемси, в урочище Сиревань – выше поселка Дубравка
- других), что может служить косвенным подтверждением расширения ареала вида. Находки Dolichophis caspius и Platyceps najadum в долинах нижнего течения рек Неожиданная, Шуюк, Джималта, окрестностях поселков Верхняя Шиловка, Голицино, Сергей-Поле, Барановка и многих других свидетельствуют о тенденции увеличения встречаемости этих видов в регионе, а находка P. najadum в 2021 году в тисовом букняке лавровишневом в среднем течении реки Восточная Хоста, на удалении более 10 км от берега моря, является прямым свидетельством расширения ареала этого средиземноморского вида.
Изменения погодных условий последнего десятилетия отражаются на фенологии и распространении рептилий. Наблюдается расширение ареалов вглубь континента и увеличение гипсометрических отметок находок у представителей средиземноморской эколого-географической группы (Testudo graeca nikolskii, Dolichophis caspius и Platyceps najadum, Natrix tessellata), а также увеличение периода активности ряда видов до декабря, включительно.
Факт современного изменения как глобального, так и регионального климата не вызывает сомнений. Климат на территории Западного Кавказа является составной частью глобальной климатической системы и его условия в рассматриваемом регионе важны своим влиянием как на сохранение биоразнообразия, так и на природопользование человека. Отмечающиеся с последней декады XX столетия изменения климата на Западном Кавказе привели к перестройкам ареалов наиболее чувствительных видов растений и животных. На Западном Кавказе в числе 9 видов ящериц имеется два узкоэндемичных вида, принадлежащих к различным биогеографическим комплексам. Первый – Darevskia derjugini – колхидский вид, распространенный в предгорьях северного макросклона Западного Кавказа в междуречье Белая – Малая Лаба и по Черноморскому побережью от горы Семашхо (Туниев, 2004, 2007) до границы с Республикой Абхазия и далее на восток до Лагодехи (Грузия) и Лазистана (Турция) на юго-востоке. Второй вид, Darevskia szczerbaki (Lukina, 1963), является угнетенным реликтом восточно-средиземноморских ценозов, до недавнего времени известный с береговых обрывов к югу от Анапы и мысов Большой и Малый Утриши. Исследования последних лет позволили сделать заключение о прогрессирующем расширении ареала Darevskia szczerbaki на более чем 120 км (Tuniyev, Tuniyev, 2012) и заметном сокращении ареала Darevskia derjugini (Туниев, 2003; Tuniyev, 2003). Предположить, что на стокилометровом участке интенсивно посещаемого побережья Черного моря ящерицы оставались просмотренными, казалось мало вероятным, и была выдвинута версия о расширении ареала Darevskia szczerbaki вдоль береговых обрывов в юго-восточном направлении. Также предполагалось, что в случае сохранения этой тенденции, Darevskia szczerbaki сможет колонизовать берег моря до Туапсе на юге, то есть на участке с подходящими биотопами, что подтвердилось позже, в связи с находкой вида в окрестностях поселка Сосновое в Туапсинском районе (Tuniyev, Tuniyev, 2012), а в 2011 году на мысе Кадош у Туапсе (Туниев, 2012а).
Обратная картина наблюдается с ареалом Darevskia derjugini. В 80–90-х годах XX века вид встречался от бассейна верхнего течения реки Аше на западе до государственной границы с Абхазией на востоке и от ущелья нижнего течения реки Сочи – горы Большой Ахун на юге до окрестностей поселка Сахрай – села Никитино на севере. Обследования последних лет показали, что вид исчез с ряда западных вершин Главного Кавказского хребта, из долины среднего течения реки Шахе и из ущелья нижнего течения реки Сочи на Черноморском побережье, а на северном склоне Западного Кавказа вид исчез из большинства периферийных
23
Туниев Б. С., Рыбак Е. А., Тимухин И. Н., Алиев Х. У.
локалитетов (Туниев, 2003; Tuniyev, 2003). Таким образом, произошло сокращение ареала Darevskia derjugini в России не менее, чем на 200 тыс. га.
По-видимому, оба наблюдаемых процесса изменения ареалов указанных видов в основе своей имеют общее происхождение. Экстремально засушливые летние сезоны 1999–2000, 2010–2012 годов негативно повлияли на всю мезофильную биоту Западного Кавказа. На кардинальную перестройку фенологических циклов живых организмов оказало влияние и общее запаздывание сезонов года, примерно на 20–25 суток, наблюдаемое с началом нового тысячелетия. К примеру, в тисо-самшитовой роще Кавказского заповедника в 2000 году произошло усыхание самшитников в возрасте более 150 лет, произрастающих в условиях маломощных почв на скальных обнажениях. Аналогичные процессы локального усыхания самшита в 1999–2000 годах наблюдались по всему Черноморскому побережью России. Не менее драматичная картина наблюдалась в 2000 году и в высокогорье: к примеру, на Лагонакском нагорье засуха приводила к образованию в почве трещин до 0,6 м глубиной. Необычно высокий и продолжительный фон летних температур последних двух десятилетий
- совокупности с продолжительным засушливым периодом отрицательно сказались на мезофильном виде – Darevskia derjugini (Туниев, 2012б) По этим же причинам, предположительно, вдоль приморской полосы Краснодарского края произошла экспансия ксерофильной восточно-средиземноморской Darevskia szczerbaki и другихвыше перечисленных восточно-средиземноморских видов рептилий.
ЗАКЛЮЧЕНИЕ
На Черноморском побережье российского Кавказа, включая территорию СНП, существующая кардинальная схема распространения средиземноморских ценозов вдоль приморской полосы соответствует сухим предгорьям с годовой суммой температур свыше 5000о и до 200–300 м н. у. м. (Tuniyev, 1995). Существование экотона двух биогеографических провинций на территории СНП в большей мере определяется изменением количества осадков, значениями минимальных (средних и абсолютных) температур, средней амплитудой относительной влажности воздуха, зоной воздействия вторжения зимних холодных северо-восточных воздушных масс (бора), в свою очередь определяемых гипсометрической высотой Главного Кавказского хребта, резко понижающегося к западу от поселка Лазаревское.
За пределами зонального Средиземноморья, на высотах до 600 м н. у. м., возрастающее количество осадков и уменьшающийся фон температуры позволяет сохраниться осколкам средиземноморских группировок внутри Колхиды исключительно в условиях локальной эдафической сухости, свойственной скалистым биотопам, термический режим которых и дефицит влажностик тому жезначительно превышают окружающие мезофитные условия на обращенных к югу и востоку крутых, преимущественно известняковых, склонах.
Исследование показало, что для средиземноморских видов приоритептное значение имеет влажность воздуха. В свете приведенных фактов изменения климатических показателей и кружева ареалов колхидских и восточно-средиземноморских видов флоры и фауны можно предположить в дальнейшем смещение к востоку условной линии, разделяющей Колхидскую и Восточно-Средиземноморскую биогеографические провинции.
Список литературы
Алешина М. А., Семенов В. А., Чернокульский А. В. Исследование роли Глобальных и региональных факторов в изменении летних осадков на Черноморском побережье Кавказа по результатам экспериментов с моделью климата // Фундаментальная и прикладная климатология. – 2019. – Т. 3. – С. 59–75.
Алиев Х. У., Туниев Б. С., Тимухин И. Н., Тания И. В. Геоботаническая и популяционная характеристики земляничника мелкоплодного (Arbutus andrachne L.) в Абхазии // Бюллетень ГНБС. – 2020. – Вып. 135. – С. 24–38.
Груза Г., Ранькова Э. Изменение климатических условий Европейской части России во второй половине ХХ века // Русский архипелаг [Электронный ресурс]. – URL http://www.archipelag.ru/agenda/geoklimat/history/change/
Джангиров М. Ю., Шевцов Б. П. Бонитетные шкалы для сосны пицундской (Pinus pityusa Steven) в Сочинском национальном парке // Известия Самарского научного центра РАН. – 2014. – Т. 16, № 5. – С. 199–204.
Доклад об особенностях климата на территории России за 2020 год. – 2021. – М. – 96 с. https://meteoinfo.ru
24
Хорологические закономерности в экотоне Восточно-Средиземноморской и Колхидской биогеографических провинций на территории Сочинского национального парка
![]()
Егошин А. В. Прогнозирование влияния климатических изменений на пространственное распределение чужеродного компонента флоры юга черноморского побережья Краснодарского края // Экосистемы. – 2021. –
- 26. – С. 23–32.
Зернов А. С. Растения Северо-Западного Закавказья. – М.: Изд-во МПГУ. – 2000. – 129 с.
Зернов А. С. Флора Северо-Западного Кавказа / [Ред. А. Г. Еленевский]. – М.: Товарищество научных изданий КМК. – 2006. – 664 с.
Изменения климата. Обобщающий доклад. Вклад рабочих групп I, II и III в четвертый доклад об оценке Межправительственной группы экспертов по изменению климата / [Ред. Пачаури Р. К., Райзингер А. и основная группа авторов]. – МГЭИК, Женева, Швейцария. – 2007. – 104 с.
Конспект флоры Кавказа: В 3 томах / [Отв. ред. Акад. А. Л. Тахтаджян]. – Т. I / [Ред. Ю. Л. Меницкий,
Т. Н. Попова]. – СПб.: Изд-во С.-Петерб. ун-та, 2003. – 204 с.
Конспект флоры Кавказа: В 3 томах / [Отв. ред. Акад. А. Л. Тахтаджян]. – Т. 2 / [Ред. Ю. Л. Меницкий,
Т. Н. Попова]. – СПб.: Изд-во С.-Петерб. ун-та, 2006. – 467 с.
Конспект флоры Кавказа: В 3 томах / [Отв. ред. Акад. А. Л. Тахтаджян]. Т. 3, ч.1 / [Ред. Ю. Л. Меницкий,
- Н. Попова, Г. Л. Кудряшова, И. В. Татанов]. – СПб.; М.: Товарищество научных изданий КМК, 2008. – 469 с. Конспект флоры Кавказа: в 3 томах / [Отв. ред. акад. А. Л. Тахтаджян]. Т. 3, ч. 2 / [Ред. Г. Л. Кудряшова,
И. В. Татанов]. – СПб.; М.: Тов-во научн. изд. КМК, 2012. – 623 с.
Кузнецов Н. И. Элементы Средиземноморской области в Западном Закавказье // Записки русского географического общества. – СПб. – 1891. – Т. 23, № 3. – 190 с.
Меницкий Ю. Л. Проект «Конспект флоры Кавказа». Карта районов флоры // Ботанический журнал. – 1991.
– Т. 76, № 11. – С. 1513–4521.
Оценочный доклад Росгидромета: Изменения климата и их последствия на территории Российской Федерации / [Ред. А. И. Бедрицкий и др.]. – Т. 1. [Электронный ресурс] URL http://www.voeikovmgo.ru/otsenochnyiy-doklad-izmenenie-klimata-na-territorii-rossiyskoy-federatsii.html
Пестерева Н. М. Комплексное исследование окружающей среды Черноморского побережья Кавказа, включая региональное изменение климата, для разработки рекомендаций устойчивого развития туристско-рекреационных кластеров исследуемого региона на период до 2020 года. – 2011. – Отчет о НИР № 5.3798.2011 от 23.11.2011 (Министерство образования и науки РФ).
Рыбак О. О., Рыбак Е. А. Чередование режимов в изменчивости североатлантического колебания // Известия ВУЗов. Естественные науки. – 2009. – № 2. – С. 60–65.
Рыбак О. О., Рыбак Е. А. Изменение температуры воздуха и количества осадков в Черноморском регионе в 20-м веке // Политематический сетевой электронный журнал Кубанского государственного аграрного университета. – 2013. – № 90. – С. 15–35. http://meteo.infospace.ru/win/ (дата обращения 20.02.2014). http://meteo.ru/pogoda-i-klimat (дата обращения 20.03.2014).
Сатунин К. А. О зоогеографических округах Кавказского края // Известия Кавказского музея. – 1912. – Т. 7, вып. 1. – С. 7–106.
Соколов С. Я. Общий естественноисторический и лесоводственный очерк Сочинского района. – Л., 1931. –
- 7–96.
Тимухин И. Н. О биогеографическом статусе горы Бозтепе Лазаревского района города Сочи // Проблемы устойчивого развития регионов рекреационной специализации. – 2008. – Сочи. – С. 205–208.
Тимухин И. Н. Высокогорная флора массива горы Аутль и её анализ // Естественные и технические науки. – 2019. –
- 11 (137). – С. 89–97.
Тимухин И. Н., Туниев Б. С. О границах Бело-Лабинского, Туапсе-Адлерского и Абхазского флористических районов Кавказа // Вестник Удмурдского университета. Биология. Науки о земле. – 2016. – Т. 26, вып. 2. – С. 91–97.
Туниев Б. С. Современная пульсация ареалов ящериц на Северо-Западном Кавказе // Актуальные вопросы экологии и охраны природы экосистем южных регионов России и сопредельных территорий: Материалы XVI межреспубликанской научно-практической конференции. – 2012а. – Краснодар: Кубанский госуниверситет. –
- 86–88.
Туниев Б. С. Первые последствия изменения климата и природопользования в биоте Западного Кавказа // Материалы Международной научной конференции Глобальные экологические процессы. – 2012б. – М.: Академия. – С. 438–443.
Туниев Б. С., Тимухин И. Н., Джангиров М. Ю. Об эксклавах средиземноморской флоры в горной Северо-Западной Колхиде // Сравнительная флористика: Анализ видового разнообразия растений. Проблемы. Перспективы. «Толмачевские чтения»: материалы X Международной школы-семинара / [Ред. С. А. Литвинская и О. Г. Баранова]. – 2014. – Краснодар: Кубанский госуниверситет. – С. 158–161.
Туниев С. Б. О распространении артвинской ящерицы – Darevskia derjugini (Nikolsky, 1898) (Reptilia: Sauria)
- Российской Федерации // Проблемы устойчивого развития регионов рекреационной специализации. – Сочи. –
2004. – С. 80–94.
Туниев С. Б. О нахождении артвинской ящерицы – Darevskia derjugini (Nikolsky, 1898) (Reptilia: Sauria) в Туапсинском районе Краснодарского края // Проблемы устойчивого развития регионов рекреационной специализации. – Сочи. – 2007. – С. 274–275.
Черепанов С. К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). – СПб.: Мир и семья — 95, 1995. – 992 с.
25
Туниев Б. С., Рыбак Е. А., Тимухин И. Н., Алиев Х. У.
![]()
Mandin J. P. Découverte de très vieux genévriers de Phénicie (Juniperus phoenicea L.) dans les gorges de l’Ardèche (France) // Le Journal De Botanique De La Société De Bontanique De France. – 2005. – Fasc. 29. – S. 53–62.
Mazur M., Minissale P., Sciandrello S., & Boratyński A. Morphological and ecological comparison of populations of Juniperus turbinata Guss. and J. phoenicea L. from the Mediterranean region // Plant Biosystems. – 2016. – Vol. 150. – P. 313–322. https://doi.org/10.1080/11263504.2014.994579
Rivas-Martínez S., Peñas Á., & Díaz González T. E. 2004. Bioclimatic Map of Europe – Thermoclimatic belts. Cartographic Service, University of León. http://www.globa lbioc limat ics.org/form/tb_med.htm
Rivas-Martínez S., Peñas Á., del Río S., Díaz T. E., & Rivas-Sáenz S. Bioclimatology of the Iberian Peninsula and the Balearic Islands // The vegetation of the Iberian Peninsula / [Ed. J. Loidi]. Plant and Vegetation. – 2017. – P. 29–80.
Rybak E. A., Rybak O. O., Zasedatelev Y. V. Complex Geographical Analysis of the Greater Sochi Region on the Black Sea Coast // GeoJournal. – 1994. – Vol. 34. – P. 507–513.
Rybak E. A., Rybak O. O. Regional effects of the global climate change; a case study: the Sochi National park area (Russia) // Nature Conservation Research. Заповедная наука. – 2017. – Т. 2, N 3. – С. 61–67.
Salvà-Catarineu M, Romo A, Mazur M. Past, present, and future geographic range of the relict Mediterranean and Macaronesian Juniperus phoenicea complex // Ecology and Evolution. – 2021. – P. 1–21. https://doi. org/10.1002/ece3.7395
Tuniyev B. S. On the Independence of the Colchis Center of Amphibian and Reptile Speciation // Asiatic Herpetological Reseaches. – 1990. – Vol. 3. – P. 67–84.
Tuniyev B. S. On the Mediterranean influence on the formation of herpetofauna of the Caucasian Isthmus and main xerophylous refugia // Russian Journal of Herpetology. – 1995. – Vol. 6, N 2. – P. 95–119.
Tuniyev B. S. Pulsation of lizard’s areas on North-West Caucasus // 12th Ordinary General Meeting Societas Europea Herpetologica. St.-Petersburg. – 2003. – P. 164.
Tuniyev B. S., Orlov N. L., Ananjeva N. B., Aghasyan A. L. Snakes of the Caucasus: taxonomic diversity,
distribution, conservation. – St. Petersburg, Moscow: KMK Scientific Press, 2019. – 276 p.
Tuniyev B. S., Tuniyev S. B. On distribution and taxonomic status of rock lizard Darevskia brauneri szczerbaki (Lukina, 1963) and D. b. darevskii (Szczerbak, 1962) // Russian Journal of Herpetology. – Vol. 19, N l. – 2012. – P. 10– 22.
Tuniyev B. S., Rybak E. A., Timukhin I. N., Aliev Kh. U. Horological patterns in the ecotone of the Eastern Mediterranean and Colchis biogeographic provinces on the territory of the Sochi National Park // Ekosistemy. 2022. Iss. 29. P. 5–26.
The article gives an overview of 11 points of study of biota and microclimate in the foothill zone of the territory of the Sochi National Park (SNP). Moreover, the researchers studied a chain of territories contrasted with the surrounding phytolandscapes with a high abundance of Mediterranean species in the SNP at a distance of up to 10 km from the sea in addition to the sub-Mediterranean coastal landscapes. The direct contact of biotas of different composition and origin, their mosaic distribution at a distance from the seacoast raises a logical question about the reasons for the formation and preservation of the ecotone of two biogeographic provinces in the relatively limited territory of the SNP. The aim of the research was to determine the features of the development of the Eastern Mediterranean and Colchis biota in the ecotone of two biogeographic eponymous chorions in the territory of the SNP from the standpoint of the microclimatic characteristics of various phyto-landscapes. The formation of the ecotone of two biogeographic provinces on the territory of the SNP is influenced by the change in the amount of precipitation, the values of the minimum (average and absolute) temperatures, the average amplitude of the relative humidity, the impact zone of the invasion of winter cold northeastern air masses (bora), which in turn are determined by hypsometric height of the Main Caucasian ridge, sharply decreasing to the west of the village of Lazarevskoye. The study showed that air humidity has the most significant impact on Mediterranean species. According to the above mentioned facts of changes in climatic indicators and the curves of the habitats of Colchis and Eastern Mediterranean species of flora and fauna, the authors assume that in the future the conditional borderline dividing the Colchis and Eastern Mediterranean biogeographic provinces will a shift to the east.
Key words: Colchis, Eastern Mediterranean, ecotone, Sochi National Park, chorology, microclimatic features.
Поступила в редакцию 23.11.21
Принята к печати 25.12.21
26
