
ЯДЕРНЫЕ АНОМАЛИИ И МИКРОЯДРА В ЭРИТРОЦИТАХ ПЕРИФЕРИЧЕСКОЙ КРОВИ КАРПОВЫХ РЫБ В РЕКАХ С ЕСТЕСТВЕННЫМ И ИЗМЕНЁННЫМ ТЕРМИЧЕСКИМ РЕЖИМОМ (РЕКИ БЕРЁЗОВКА И ТЁПЛАЯ, БАССЕЙН РЕКИ ЕНИСЕЙ)
NUCLEAR ANOMALIES AND MICRONUCLEI IN PERIPHERAL BLOOD ERYTHROCYTES OF CYPRINID FISH IN RIVERS WITH NATURAL AND ALTERED THERMAL REGIMES (BERYOZOVKA AND TEPLAYA RIVERS, YENISEI RIVER BASIN)
JOURNAL: «EKOSISTEMY», Issue 41, 2025
Publication text (PDF):Download
UDK: 597.554; 591.11
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Yablokov N. O.
Research Institute of Ecology of Fishery Reservoirs. Krasnoyarsk Branch of the All-Russian Research Institute of Fisheries and Oceanography, Krasnoyarsk, Russia
TYPE: Article
DOI: https://doi.org/10.29039/2413-1733-2025-41-147-157
PAGES: from 147 to 157
STATUS: Published
LANGUAGE: Russian
KEYWORDS: cyprinid fishes, blood, micronuclei, erythrocyte anomalies, thermal pollution, Teplaya River, Beryozovka River.
ABSTRACT (ENGLISH):
The paper presents data on the occurrence of micronuclei and nuclear anomalies in peripheral blood erythrocytes of three carp species: the gudgeon Gobio gobio (Linnaeus, 1759), the Eurasian minnow Phoxinus phoxinus (Linnaeus, 1758), and the sunbleak Leucaspius delineatus (Heckel, 1843)). These species inhabit rivers with both natural (Beryozovka River) and altered thermal regimes (Teplaya River). During the study, an extensive list of nuclear anomalies – 20 and 22 types, respectively – was found in the erythrocytes of fish from both the Teplaya and Beryozovka Rivers. The occurrence of abnormal nuclei in erythrocytes of the three studied fish species from the Teplaya River averaged 1.52±0.25–2.61±0.91 %, while in fish from the Beryozovka River, it ranged from 2.27±0,63–3.53±0.97 %. The assessment of the significance of differences between the occurrences of anomalies revealed no statistically significant differences among individuals of each of the fish species from the two studied watercourses. The values of the Mann-Whitney test (U) for the gudgeon samples were 74.0 (p=0.45), for the Eurasian minnow were 39.0 (p=0.43), and for the sunbleak were 29.0 (p= 0.96). Despite the absence of statistically significant differences, variations in the occurrence of specific types of anomalies were observed among the erythrocyte samples. Based on the obtained data, an assumption was made that the intensity of anomalies in the erythrocyte nuclei of carp fish living in the Teplaya River was influenced more by hydrochemical factors than by the altered thermal regime.
ВВЕДЕНИЕ
Термическое загрязнение водных объектов, вызываемое сбросом подогретых вод промышленных предприятий, тепловых и атомных электростанций, является одной из серьезнейших экологических проблем современности (Verones et al., 2010; Raptis et al., 2016). Повышение температуры естественных водоёмов и водотоков, за счет поступления тёплых вод, оказывает воздействие на гидрохимический режим водоёма (снижение количества растворённого кислорода, увеличения скорости химических реакций, повышение трофности), что в совокупности приводит к структурным и количественным изменениям в составе сообществ гидробионтов (Verones et al., 2010; Шихмарёв, 2013).
Одним из структурных компонентов водных экосистем, испытывающих значительное воздействие термического загрязнения, является рыбное сообщество. Температура воды оказывает влияние на скорость основных биологических процессов рыб (рост, питание, обмен веществ и т.д.), а также их поведение и пространственное распределение (Голованов, 2013). Значительные отклонения температуры за пределы оптимальных величин являются стресс-факторами, снижающими адаптационные способности организма. В частности, продолжительное воздействие повышенной температуры воды может приводить к смещению сроков нереста, фаз жизненного цикла и более раннему созреванию половых продуктов (Голованов и др., 2005; Шихмарёв, 2013). Наряду с температурой, воздействие на организм рыб в местах сброса подогретых вод, может оказывать химическое загрязнение, поскольку повышенные значения температуры воды могут усиливать действие некоторых токсических веществ (нефтепродуктов, хлорорганических соединений, тяжелых металлов и др.) (Голованов, 2013). Комплекс перечисленных факторов может служить причиной возникновения спонтанных мутаций, приводящих к нарушениям стабильности генома и эффективной передачи генетического материала (Sylvester, 1975; Hassan et al., 2017).
В бассейне реки Енисей одним из примеров водного объекта, испытывающего термическое загрязнение, служит река Тёплая, расположенная в черте города Красноярска. Этот левобережный приток реки Берёзовка (правый приток реки Енисей), длиной около трёх километров, на значительном протяжении представляет собой сбросной канал, принимающий подогретые воды Красноярской ТЭЦ-1. Кроме того, по данным региональных средств массовой информации, в акватории реки регистрируются случаи сброса неочищенных промышленных и ливневых стоков. Учитывая тот факт, что река Тёплая, как и река Берёзовка, являются популярными местами любительского рыболовства среди жителей города Красноярска и посёлка Берёзовка, оценка ихтиоценозов рассматриваемых водотоков имеет значение с точки зрения сохранения и рационального использования водных биологических ресурсов.
В настоящее время, при оценке мутагенной активности факторов антропогенного воздействия на живые организмы широкое применение получили методы микроядерного тестирования и оценки встречаемости морфологических аномалий клеточных ядер (Ayllon, Garcia-Vazquez, 2000; Ильинских и др., 2011; Hayashi, 2016; Arellano-García, 2021). Наличие ядер в эритроцитах периферической крови рыб делает эти клетки удобным объектом для изучения влияния факторов среды на стабильность генетического материала (Крюков, 2023).
Цель данной работы – оценить влияние термического загрязнения реки Тёплая на популяции массовых видов рыб, обитающих в указанном водном объекте, на основании показателей встречаемости микроядер и ядерных аномалий в эритроцитах их периферической крови.
МАТЕРИАЛ И МЕТОДЫ
Объектами исследования послужили три вида рыб (пескарь Gobio gobio (Linnaeus, 1759), гольян речной Phoxinus phoxinus (Linnaeus, 1758) и верховка Leucaspius delineatus (Heckel, 1843)), являющиеся наиболее массовыми и широко распространёнными видами рыб в системе реки Берёзовка. Ихтиологический материал отбирали в июне–августе 2024 года в р. Тёплая (56°02′47.65″ с.ш., 93°06′17.88″ в.д.) и в нижнем течении реки Берёзовка, выше впадения Тёплой (56°02′12.50″ с.ш., 93°06′59.67″ в.д.). В качестве орудия лова использован гидробиологический сачок. У отловленных особей в соответствии с общепринятыми ихтиологическими методиками измеряли абсолютную длину и массу тела (Правдин, 1966). Сводная информация об объёме выборки рыб, взятых на анализ, и их размерных показателях представлена в таблице 1.
После проведения измерений длины и массы рыб выполняли забор крови. Кровь отбирали из хвостовой артерии путем отсечения хвостового стебля у анального плавника. Мазки подготавливали непосредственно после отлова рыб и после высыхания фиксировали 96 % этиловым спиртом. Последующее окрашивание мазков для определения количества микроядер и аномалий ядер эритроцитов проводили по методу Романовского–Гимзы в лабораторных условиях (Житенева и др., 2004). Окрашенные препараты анализировали на микроскопе БЛМ-М1, оснащенном цифровой камерой ToupCam 14 MP. В каждом препарате просматривали не менее 500 эритроцитов, затем вычисляли частоту встречаемости эритроцитов с микроядрами (отношение числа клеток с микроядрами к общему числу просмотренных клеток, в %). Аналогичным образом вычисляли встречаемость аномалий ядер эритроцитов. Алгоритм выявления аномалий и их протоколирование выполнены в соответствии с работой В. И. Крюкова (2023).
Таблица 1
Абсолютная длина и масса тела исследованных рыб из рек Тёплая и Берёзовка
| Вид рыбы | Абсолютная длина, мм | Масса тела, г | Число экземпляров |
| река Тёплая | |||
| Гольян речной | 38,8±2,729,0–55,0 | 0,85±0,230,22–2,21 | 10 |
| Пескарь | 48,8±6,730,0–110,0 | 2,42±1,060,32–12,85 | 12 |
| Верховка | 50,4±3,734,0–68,0 | 1,26±0,240,32–2,37 | 12 |
| река Берёзовка | |||
| Гольян речной | 36,1±0,732,0–39,0 | 0,38±0,020,23–0,47 | 10 |
| Пескарь | 38,9±1,830,0–52,0 | 0,51±0,071,12–6,06 | 15 |
| Верховка | 40,4±0,639,0–42,0 | 0,57±0,050,43–0,43 | 5 |
Примечание к таблице. Среднее значение (m) ± стандартная ошибка (SE), под чертой – минимальные и максимальные значения.
Для выявления различий в температурных режимах рек Тёплая и Берёзовка в местах отбора ихтиологического материала в период с апреля по ноябрь термооксиметром МАРК 302Э с периодичностью два раза в месяц проводили измерения температуры воды (с точностью измерений 0,1 ºС).
Статистический анализ данных проведен методами описательной статистики. Данные о размерах рыб и встречаемости аномальных ядер представлены как среднее значение и его стандартная ошибка. Доли отдельных типов аномалий представлены как процент (%) от общего числа проанализированных клеток в выборке. Достоверность различий между выборками оценивали по непараметрическому критерию Манна-Уитни (U-критерий). Для выявления различий в процентном составе отдельных типов ядерных аномалий в эритроцитах карповых рыб из рек Берёзовка и Тёплая полученные данные (% от общего числа проанализированных клеток в выборке) оценивались посредством мультивариантного анализа методом главных компонент. Результаты анализа представлены в виде диаграмм рассеяния отдельных проб рыб в пространстве двух первых главных компонент (ГК1 и ГК2) и таблицы факторных нагрузок по каждому из обнаруженных типов аномалий.
Статистическая обработка данных выполнена в программных пакетах Statistica 10 (StatSoft Inc., США) и PAST 4.02 (Осло, Норвегия). Построение диаграмм рассеяния при проведении мультивариантного анализа методом главных компонент выполнено в графической онлайн-платформе SRplot (Tang et al., 2023).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В ходе проведённого анализа крови у каждой из исследованных рыб были обнаружены аномалии ядер эритроцитов. Общий перечень аномалий, обнаруженный в мазках крови рыб из рек Берёзовка и Тёплая включал 22 типа аномалий ядер эритроцитов, в т.ч.: микроядра (обособленные и примыкающие), двуядерные клетки (с изолированными ядрами, с примыкающими ядрами, с гантелевидными ядрами и с ядрами-«восьмерками), двулопастные ядра, почкующиеся ядра, пузырящиеся ядра, хвостатые ядра с клювовидным отростком, зазубренные ядра (клинообразная инвагинация ядерной оболочки), ядра с впадиной
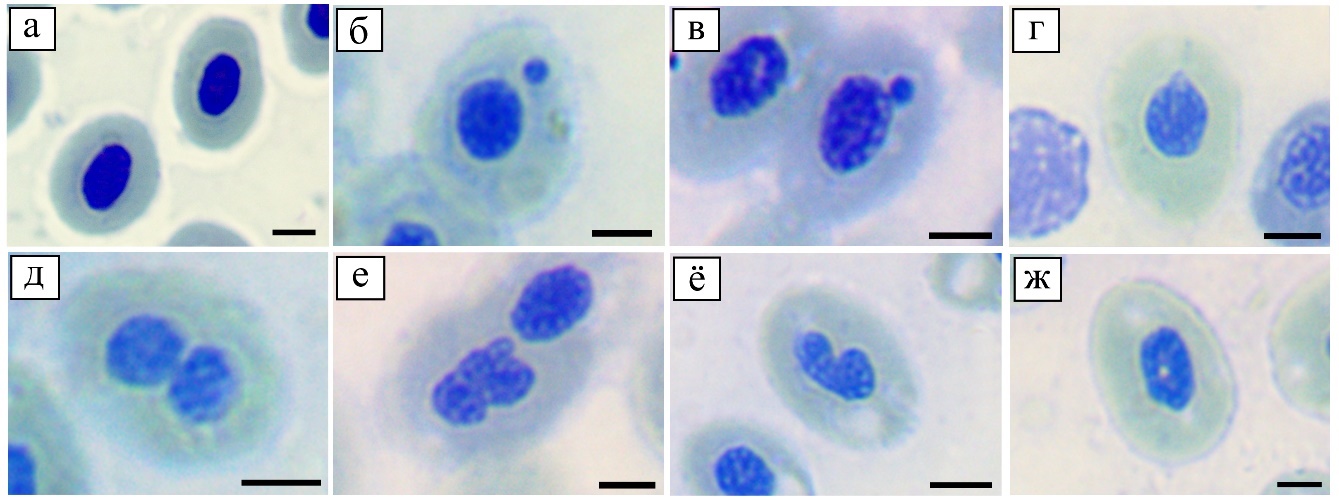
Рис. 1. Нормальные и аномальные ядра эритроцитов, обнаруженные у рыб
из рек Тёплая и Берёзовка
а – эритроциты с нормальными ядрами (верховка, река Берёзовка), б – изолированное микроядро (гольян, река Берёзовка), в – примыкающее микроядро (пескарь, река Тёплая), г – хвостатое ядро с клювовидным отростком (гольян, река Берёзовка), д – двулопастное ядро (гольян, река Берёзовка), е – почкующееся ядро (гольян, река Берёзовка), ё – зазубренное ядро (гольян, река Тёплая), ж – ядро с вакуолью (гольян, река Берёзовка). Размер мерной линейки – 5 мкм.
(инвагинация ядерной оболочки с пологими скатами), почковидные ядра, вакуолизированные ядра, кариопикноз, кариорексис, кариолизис, клетки со смещёнными ядрами, микроциты, а также безъядерные клетки (рис. 1).
В эритроцитах периферической крови рыб из реки Берёзовка зарегистрированы все вышеперечисленные типы аномалий. При этом у речного гольяна и пескаря было отмечено по 16 типов аномалий, у верховки – 17 типов. Преобладающим по частоте встречаемости типом аномалий для всех изученных рыб выступали эритроциты со смещенными ядрами (от 0,83 % от общего числа исследованных клеток у речного гольяна до 2,25 % у верховки). Субдоминирующими типами аномалий выступали ядра с впадиной (у речного гольяна и верховки) и микроциты (у пескаря) (табл. 2).
В эритроцитах рыб из реки Тёплая отмечено 20 типов аномалий, в т.ч. у гольяна и верховки – по 13 типов, у пескаря – 12. Среди отмеченных аномалий у пескаря и верховки количественно преобладали эритроциты со смещёнными ядрами (1,46 и 1,74 % соответственно), у гольяна – ядра с впадиной (0,68 %). У гольянов из реки Тёплая преобладающим типом аномалий были ядра с впадиной. Стоит отметить, что у значительного числа рыб как из реки Тёплая, так и из реки Берёзовка, обилие смещённых ядер было сопряжено с обширным пойкилоцитозом клеток. К субдоминрующим типам аномалий у пескаря отнесены микроциты (0,21 %), у верховки – кариорексис (0,12 %), у речного гольяна – клетки со смещёнными ядрами (0,41 %) (см. табл. 2).
Встречаемость аномальных ядер в эритроцитах рыб из реки Берёзовка составила 2,27±0,63 % у гольяна, 3,53±0,97 % у пескаря и 2,97±1,28 % у верховки. У рыб из реки Тёплая – 1,52±0,25, 2,61±0,91 и 2,40±0,44 % соответственно. Оценка значимости различий между встречаемостью аномалий, выполненная посредством критерия Манна-Уитни (U), показала отсутствие статистически значимых различий между особями каждого из исследованных видов рыб из рек Тёплая и Берёзовка. Значения U-критерия для выборок пескаря составили 74,0 (при p=0,45), для речного гольяна – 39,0 (при p=0,43), для верховки – 29,0 (при p=0,96).
Несмотря на отсутствие достоверных различий, между выборками по общей встречаемости аномалий эритроцитов в выборках из рек с естественным и изменённым термическим режимами наблюдались различия по встречаемости отдельных типов аномалий (табл. 2, 3, рис. 2).
Таблица 2
Встречаемость различных типов аномалий (% от общего числа проанализированных клеток в выборке) в эритроцитах карповых рыб из рек Берёзовка и Тёплая
| № | Тип аномалии в эритроците | Гольян | Пескарь | Верховка | ||||
| Тёплая (n = 10) | Берёзовка (n = 10) | Тёплая (n = 12) | Бёрезовка (n = 15) | Тёплая (n =12) | Бёрезовка (n = 5) | |||
| 1 | Микроядра изолированные | 0,036 | 0,027 | 0,068 | 0,085 | 0,006 | 0,013 | |
| 2 | Микроядра примыкающие | – | 0,020 | 0,038 | 0,011 | 0,013 | 0,013 | |
| 3 | Двуядерные клетки с изолированными ядрами | – | 0,040 | 0,008 | 0,006 | – | – | |
| 4 | Двуядерные клетки с примыкающими ядрами | 0,012 | 0,087 | – | – | – | 0,013 | |
| 5 | Двуядерные клетки с гантелевидными ядрами | – | 0,007 | – | 0,006 | – | – | |
| 6 | Двуядерные клетки с ядрами-«восьмёрками» | 0,006 | 0,013 | 0,008 | – | – | 0,027 | |
| 7 | Двулопастные ядра | 0,006 | – | – | 0,017 | 0,019 | 0,013 | |
| 8 | Лопастное ядро с лопастью соединённой мостом | – | – | – | – | 0,006 | – | |
| 9 | Лопастные ядра с примыкающей лопастью | – | – | – | – | – | 0,027 | |
| 10 | Почкующиеся ядра | 0,053 | 0,033 | 0,015 | 0,068 | 0,013 | 0,013 | |
| 11 | Пузырящиеся ядра | 0,059 | 0,027 | 0,190 | 0,040 | – | 0,013 | |
| 12 | Хвостатые ядра с клювовидным отростком | – | 0,033 | – | 0,023 | 0,031 | – | |
| 13 | Зазубренные ядра | 0,124 | 0,220 | 0,008 | 0,011 | 0,031 | 0,066 | |
| 14 | Ядра с впадиной | 0,675 | 0,553 | 0,023 | 0,023 | 0,063 | 0,120 | |
| 15 | Почковидные ядра | 0,142 | 0,053 | 0,030 | 0,017 | – | – | |
| 16 | Вакуолизированные ядра | 0,006 | 0,053 | – | 0,006 | – | – | |
| 17 | Кариопикноз | 0,024 | 0,047 | 0,068 | 0,124 | 0,069 | 0,027 | |
| 18 | Кариорексис | – | – | – | 0,017 | 0,239 | 0,013 | |
| 19 | Кариолизис | – | – | – | – | 0,069 | 0,040 | |
| 20 | Смещённые ядра | 0,414 | 0,826 | 1,460 | 2,924 | 1,737 | 2,246 | |
| 21 | Микроциты | 0,018 | 0,193 | 0,479 | 0,209 | 0,057 | 0,027 | |
| 22 | Безъядерные эритроциты | – | – | – | – | – | 0,016 | |
| 23 | Эритроциты с нормальными ядрами | 98,426 | 97,769 | 97,604 | 96,415 | 97,646 | 97,315 | |
| Всего проанализированных клеток | 16900 | 15015 | 13148 | 17714 | 15891 | 7524 | ||
| Встречаемость аномальных ядер, % | 1,52±0,250,65–2,74 | 2,27±0,630,44–7,12 | 2,61±0,910,72–12,36 | 3,53±0,970,22–12,72 | 2,40±0,440,60–5,82 | 2,97±1,280,50–7,65 | ||
При рассмотрении данного показателя, применительно к отдельным видам рыб, наибольший уровень дисперсии отмечен для выборок речного гольяна (общая дисперсия по двум главным компонентам – 38,5 %), при этом различия проявлялись в обилии эритроцитов со смещёнными и зазубренными ядрами, а также ядрами, подверженными кариолизису, у рыб из реки Берёзовка; а также ядрами с впадиной и почковидными ядрами у рыб из реки Тёплая. Показатели общей дисперсии (в пространстве двух главных компонент) для пескаря и верховки составили 31,6 и 33,6 % соответственно. Факторные коэффициенты отдельных типов ядерных аномалий в эритроцитах карповых рыб из рек Берёзовка и Тёплая представлены в таблице 3.
Анализ данных о частоте встречаемости аномалий эритроцитов у трёх видов рыб с использованием метода главных компонент выявил, что наблюдаемые различия в
Таблица 3
Факторные коэффициенты отдельных типов ядерных аномалий
в эритроцитах карповых рыб из рек Берёзовка и Тёплая
| Тип аномалии | Гольян | Пескарь | Верховка | Все виды | ||||
| ГК1 | ГК2 | ГК1 | ГК2 | ГК1 | ГК2 | ГК1 | ГК2 | |
| Микроядра изолированные | -0,140 | -0,286 | -0,448 | 0,002 | -0,129 | 0,436 | -0,034 | 0,342 |
| Микроядра примыкающие | 0,117 | -0,163 | -0,079 | 0,113 | 0,235 | 0,246 | 0,079 | 0,191 |
| Двуядерные клетки с изолированными ядрами | – | – | 0,065 | -0,073 | – | – | 0,057 | 0,017 |
| Двуядерные клетки с примыкающими ядрами | 0,215 | 0,233 | – | – | 0,114 | 0,097 | -0,088 | 0,019 |
| Двуядерные клетки с гантелевидными ядрами | – | – | -0,040 | 0,034 | – | – | 0,059 | 0,170 |
| Двуядерные клетки с ядрами-«восьмёрками» | -0,224 | 0,421 | -0,085 | 0,108 | -0,419 | -0,055 | -0,053 | -0,351 |
| Двулопастные ядра | 0,050 | -0,156 | 0,052 | -0,580 | -0,281 | 0,121 | 0,028 | -0,105 |
| Лопастное ядро с лопастью соединённой мостом | – | – | – | – | 0,267 | 0,097 | 0,073 | 0,077 |
| Лопастные ядра с примыкающей лопастью | – | – | – | – | -0,341 | 0,144 | 0,094 | -0,262 |
| Почкующиеся ядра | 0,008 | 0,015 | -0,409 | -0,066 | 0,294 | 0,136 | -0,048 | 0,288 |
| Пузырящиеся ядра | 0,186 | 0,191 | -0,208 | 0,083 | 0,101 | -0,125 | 0,001 | 0,280 |
| Хвостатые ядра с клювовидным отростком | -0,372 | -0,151 | 0,052 | 0,071 | -0,088 | 0,318 | -0,177 | 0,011 |
| Зазубренные ядра | -0,407 | -0,144 | -0,352 | -0,146 | -0,125 | -0,279 | -0,466 | -0,136 |
| Ядра с впадиной | -0,440 | 0,278 | -0,454 | 0,054 | -0,259 | 0,365 | -0,542 | -0,104 |
| Почковидные ядра | -0,385 | 0,226 | -0,057 | -0,482 | – | – | -0,460 | -0,059 |
| Вакуолизированные ядра | -0,206 | -0,365 | 0,039 | -0,571 | – | – | -0,282 | -0,015 |
| Кариопикноз | -0,226 | 0,412 | -0,333 | -0,096 | 0,242 | -0,210 | 0,039 | 0,161 |
| Кариорексис | – | – | 0,137 | 0,065 | 0,039 | -0,324 | 0,211 | -0,325 |
| Кариолизис | – | – | – | – | -0,190 | -0,401 | 0,213 | -0,437 |
| Смещённые ядра | 0,110 | 0,119 | – | – | -0,244 | -0,176 | 0,185 | 0,025 |
| Микроциты | -0,286 | -0,331 | -0,275 | 0,027 | 0,274 | 0,045 | -0,001 | 0,171 |
| Безъядерные эритроциты | – | – | 0,159 | -0,143 | -0,235 | -0,059 | -0,023 | -0,249 |
Примечание к таблице. ГК1, ГК2 – главные компоненты.
пространстве первых двух компонент, которые вместе объясняют 22,2 % дисперсии, указывают на то, что основная часть вариаций частоты различных типов аномалий обусловлена межвидовыми отличиями, а не воздействием подогретых вод (рис. 2).
Существующие литературные данные о воздействии повышенной температуры воды на встречаемость микроядер и аномалий в ядрах эритроцитов периферической крови рыб противоречивы, что, по всей видимости, объясняется отсутствием единой методики типирования и количественной оценки аномалий эритроцитов, а также различными температурными критериями жизнедеятельности исследуемых рыб. Влияние подогретых вод в качестве одного из факторов, повышающих вероятность образования микроядер в крови рыб (плотвы Rutilus rutilus Linnaeus, 1758 и леща Abramis brama Linnaeus, 1758) из загрязнённых водоёмов (на примере Рыбинского и Горьковского водохранилищ) рассматривалось Ю. Г. Изюмовым с соавторами (2003). Увеличение частоты встречаемости микроцитов и повережденных эритроцитов в периферической крови серебряного карася отмечено в эксперименте при повышении температуры воды до критического термического
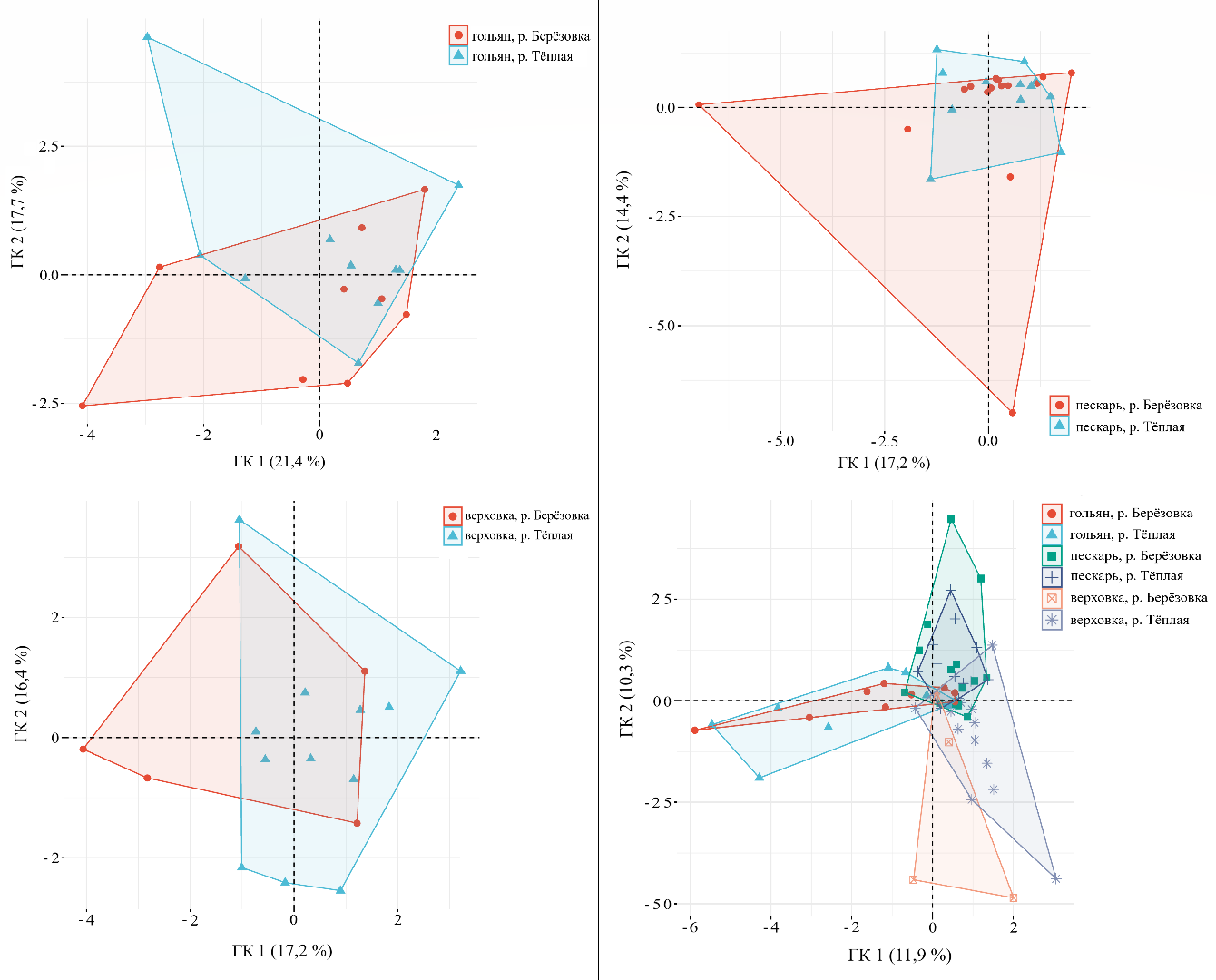
Рис. 2. Мультивариантный анализ методом главных компонент (ГК) встречаемости типов ядерных аномалий (% от общего числа проанализированных клеток в выборке) в эритроцитах карповых рыб из рек Берёзовка и Тёплая
Индивидуальные переменные – пробы рыб.
максимума (Голованов и др., 2016). В работе Т. А. Шеиной с соавторами (2022) выявлено комплексное влияние повышения минерализации вод и температуры на встречаемость некоторых типов аномалий клеток крови (пойкилоцитоз, ацентрическое расположение ядра, фестончатый контур, вакуолизация цитоплазмы, ядерные тени) речного окуня. В то же время, согласно работе Т. А. Кухаревой и Е. С. Кладченко (2021) гипертермия (впрочем, как и гипотермия), не влияла на число аномалий эритроцитов у бычка-кругляка Neogobius melanostomus (Pallas, 1814).
Стоит отметить, что эколого-физиологические температурные оптимумы для пескаря и верховки составляют около 20 °С (Голованов, 2013; Голованов, Капшай, 2015), то есть температура воды из реки Берёзовка (по данным измерений 2024 года) не превышает оптимальных значений температуры воды даже в летний период, а из реки Тёплая – лишь незначительно на 3–4 °С. Несколько ниже величина температурного оптимума у речного гольяна – 16,8 °С. Температура воды из реки Тёплая превышала значения температурного оптимума для речного гольяна с начала мая до конца августа, в то время как в реке Берёзовка – лишь непродолжительный промежуток времени (рис. 3). Возможно, именно по этой причине показатели / аномалии крови гольяна характеризовались наибольшей дисперсией при проведении мультивариантного анализа методом главных компонент.
Рис. 3. Сезонная динамика температуры воды в реке Тёплая и устьевой части реки Берёзовка (апрель – ноябрь 2024 года)
Ввиду отсутствия общепринятой шкалы референсных значений, позволяющей оценить благополучие состояния популяций рыб в водном объекте по общим показателям встречаемости аномалий ядер эритроцитов, оценка состояния популяций рыб в реках Тёплая и Берёзовка на основании данного показателя крайне затруднительна (Крюков, 2023). Однако стоит отметить, что полученные значения суммарной частоты встречаемости всех изученных типов аномалий ядер эритроцитов у рыб из рек Тёплая и Берёзовка (1,52–3,53 %) значительно превышают фоновые значения, экспериментально установленные для некоторых видов рыб (Есин, Нуриахметов, 2018; Крюков и др., 2020). Так, например, у контрольных групп данио-рерио Danio rerio (Hamilton, 1822) встречаемость ядерных аномалий эритроцитов составляет около 0,15–0,18 % (Есин, Нуриахметов, 2018; Крюков и др., 2020), у сазана Cyprinus carpio Linnaeus, 1758 – 0,75 % (Крюков, 2018; 2019)
Среди исследованных в работе типов аберраций ядер эритроцитов наиболее часто для оценки генотоксичного воздействия факторов среды применяются показатели встречаемости микроядер (Ильинских и др., 2011; Крюков, 2023). У исследованных в настоящей работе рыб средняя частота встречаемости клеток с микроядрами находилась в пределах допустимых значений, известных по работе Н. Н. Ильинских с соавторами (2011), согласно которым в норме встречаемость клеток с микроядрами при спонтанном мутагенезе не превышает 0,05–0,10 %. Лишь у пескаря из реки Тёплой величина этого показателя незначительно превышала допустимый диапазон (0,11 %). Следует отметить, что частота встречаемости микроядер в эритроцитах рыб в исследованных водных объектах была близка к средним значениям данного показателя у рыб, обитающих в нижнем течении реки Енисей (0,04–0,10 %) (Крюков, Кочкарёв, 2013).
При сравнении показателей встречаемости эритроцитов со смещённым ядром (наиболее распространённого типа аномалий у исследуемых рыб) с данными о встречаемости этого типа аномалий у некоторых карповых рыб, населяющих бассейн Волги (Минеев, 2016; Минеев, Минеева, 2019; Минеев, 2020), отмечено, что указанный тип аномалий преобладал также у большинства рыб из представленных в литературе выборок. При этом у пескаря из условно чистого водного объекта (пруд Гранный) встречаемость эритроцитов со смещенными ядрами составляла 1,29 % (Минеев, Минеева, 2019), у уклейки Alburnus alburnus (Linnaeus, 1758) и плотвы из слабозагрязнённой реки Большой Черемшан – 1,72 и 3,75 % соответственно (Минеев, 2020), а у плотвы и леща из загрязнённого Куйбышевского водохранилища повышалась до 11–12 % (Минеев, 2016).
На основании полученных данных можно предположить, что на интенсивность возникновения аномалий в ядрах эритроцитов карповых рыб, обитающих в реках Тёплая и Берёзовка, в большей степени оказывал воздействие не изменённый термический режим, а иные неблагоприятные факторы среды, среди которых наиболее вероятным является загрязнённость вод.
Несмотря на то, что регулярная оценка качества вод в нижнем течении реки Берёзовка не проводится, имеются отдельные работы, посвященные оценке качества вод по гидробиологическим показателям (Кислицина, 2019; Никонорова, Лукашина, 2020). По имеющимся данным, степень загрязнённости воды в реке Берёзовка (в нижнем течении) по показателям зообентоса соответствует II–III классам качества и степенью загрязненности вод «слабо загрязнённая» – «загрязнённая». Кроме того, ежегодные мониторинговые исследования качества вод по гидрохимическим показателям проводятся в верховьях реки (река Малая Берёзовка). По данным Государственного доклада «О состоянии и охране окружающей среды в Красноярском крае в 2023 году» (2024) по повторяемости случаев превышения ПДК загрязнённость воды в реке Малая Берёзовка по показателям: азот нитритный, запах, фосфор фосфатный, характеризуется как «неустойчивая»; по показателям: алюминий, железо общее, ХПК – как «характерная»; по показателям: БПК5, марганец, медь, фенолы летучие, цинк – как «устойчивая». Сведения о гидрохимических характеристиках воды из реки Тёплая в настоящее время отсутствуют, однако по косвенным признакам (расположение в водосборе реки промышленных предприятий, захламлённость поймы реки бытовыми отходами, периодические случаи сброса неочищенных промышленных и ливневых стоков) можно судить о загрязнённости данного водного объекта.
ЗАКЛЮЧЕНИЕ
В ходе работы проведён анализ разнообразия и встречаемости ядерных аномалий и микроцитов в периферической крови трёх карповых рыб (гольян речной, пескарь, верховка), обитающих в реках с естественным (река Берёзовка) и изменённым термическим режимом (река Тёплая).
В эритроцитах рыб, населяющих как реку Тёплая, так и реку Берёзовка, обнаружен обширный перечень ядерных аномалий (20 и 22 типа аномалий соответственно). Встречаемость аномальных ядер в эритроцитах рыб из реки Тёплая составила в среднем от 1,52±0,25 до 2,61±0,91 %, у рыб из реки Берёзовка – от 2,27±0,63 до 3,53±0,97 %. Преобладающим по частоте встречаемости типом аномалий для большинства изученных рыб выступали эритроциты со смещенными ядрами. У гольянов из реки Тёплая количественно преобладали ядра с впадиной. Достоверные различия по частоте встречаемости аномалий ядер эритроцитов между особями каждого из исследованных видов рыб из рек Тёплая и Берёзовка не выявлены. Различия обнаружены исключительно по частоте встречаемости отдельных типов аномалий.
Предположено, что на интенсивность возникновения аномалий в ядрах эритроцитов карповых рыб, обитающих в реках Тёплая и Берёзовка, в большей степени оказывали воздействие гидрохимические факторы, нежели изменённый термический режим. Для оценки уровня воздействия факторов среды на состояние ихтиофауны указанных водных объектов возникает необходимость дальнейших комплексных исследований, включающих анализ, как гидрохимических показателей среды, так и цитоморфологических исследований клеток периферической крови рыб.
Благодарности. Автор выражает благодарность ведущему специалисту лаборатории гидробиологии Красноярского филиала ГНЦ РФ ФГБНУ «ВНИРО» («НИИЭРВ») Кислициной Н.И. за помощь в проведении мультивариантного анализа методом главных компонент.
- Голованов В. К. Температурные критерии жизнедеятельности пресноводных рыб. – М.: ПОЛИГРАФ-ПЛЮС, 2013. – 300 с
- Голованов В. К., Капшай Д. С. Сравнительный анализ температурного оптимума и верхней температурной границы жизнедеятельности у молоди рыб, обитающих в водоемах Верхней Волги // Труды Института биологии внутренних вод РАН. – 2015. – № 72 (75). – С. 80–90.
- Голованов В. К., Заботкина Е. А., Некрутов Н. С., Грачёва Е. Л. Влияние высокой температуры на показатели крови у молоди серебряного карася Carassius auratus и головешки-ротана Perccottus glenii // Вестник Астраханского государственного технического университета. Серия: Рыбное хозяйство. – 2016. – №. 1. – С. 99–106.
- Государственный доклад «О состоянии и охране окружающей среды в Красноярском крае в 2023 году». – Красноярск, 2024. – 358 с.
- Есин Т. А., Нуриахметов А. А. Фоновый уровень ядерных аномалий в эритроцитах рыбы Danio rerio // Известия Российской Военно-медицинской академии. – 2018. – Т. 37, № 1. – С. 214–217.
- Житенева Л. Д., Макаров Э. В., Рудницкая О. А. Основы ихтиогематологии (в сравнительном аспекте). –Ростов-на-Дону: Эверест, 2004. – 311 с.
- Изюмов Ю. Г., Таликина М. Г., Чеботарёва Ю. В. Количество микроядер в эритроцитах периферической крови плотвы Rutilus rutilus (L.) и леща Abramis brama (L.) Рыбинского и Горьковского водохранилищ // Биология внутренних вод. – 2003. – № 1. – С. 98–101.
- Ильинских Н. Н., Ксенц А. С., Ильинских Е. Н, Васильев С. А., Манских В. Н., Ильинских И. Н. Микроядерный анализ в оценке цитогенетической нестабильности. – Томск: Изд-во ТГПУ, 2011. – 234 с.
- Кислицина Н. И. Структура зообентоса приустьевых районов рек Берёзовка и Есауловка (Красноярский край). – Красноярск: Сибирский федеральный университет, 2019. – 64 с.
- Крюков В. И. Индукция микроядер в эритроцитах карпа при сочетанном действии ионов меди и низкочастотного переменного электромагнитного поля // Биология в сельском хозяйстве. – 2018. – Т. 19, № 2. – С. 17–31.
- Крюков В. И. Анализ микроядер и ядерных аномалий в эритроцитах рыб, амфибий, рептилий и птиц: критерии выявления и типирования. – Красноярск: Научно-инновационный центр, 2023. – 94 с.
- Крюков В. И. Влияние ионов кадмия на индукцию ядерных аномалий в эритроцитах карпов // Биология в сельском хозяйстве. – 2019. – Т. 22, № 1. – С. 11–17.
- Крюков В. И., Лактюшина Н. В., Беляева А. О. Индукция микроядер в эритроцитах рыб водными вытяжками из ванадийсодержащих промотходов, захороненных на территории орловской городской свалки ТБО // Биология в сельском хозяйстве. – 2020. – Т. 27, № 2. – С. 2–12.
- Кухарева Т. А., Кладченко Е. С. Влияние факторов водной среды и физиологических состояний организма на количество аномальных эритроцитов у донных рыб Черного моря // Комплексные исследования Мирового океана. Материалы VI Всероссийской научной конференции молодых ученых (Москва, 18–24 апреля 2021 года). – Москва: Институт океанологии им. П. П. Ширшова РАН, 2021. – С. 272–273.
- Минеев А. К. Гематопатологии у рыб Куйбышевского водохранилища // Известия Самарского научного центра Российской академии наук. – 2016. – Т. 18, № 5–1. – С. 51–59.
- Минеев А. К., Минеева О. В. Гематологические параметры и паразитофауна обыкновенного пескаря Gobio gobio (Linnaeus, 1758) // Вестник Томского государственного университета. Биология. – 2019. – № 47. – С. 123–141.
- Минеев А. К. Патологии эритроцитов у рыб р. Большой Черемшан // Экологический сборник 7: Труды молодых ученых. Всероссийская (с международным участием) молодежная научная конференция. – Учреждение Российской академии наук Институт экологии Волжского бассейна РАН, 2019. – № 1. – С. 316–318.
- Никонорова Д. В., Лукашина Л. С. Оценка экологического состояния реки Березовка (Красноярский край) по гидробиологическим показателям // Сборник материалов V Всероссийской научно-практической конференции магистрантов (Москва, 16–17 апреля 2020 года). Часть 2. – Москва: ООО «Русайнс», 2020. – С. 353–360.
- Правдин И. Ф. Руководство по изучению рыб (преимущественно пресноводных). – М.: Пищевая промышленность, 1966. – 376 с.
- Шеина Т. А., Михеев П. Б., Мухина М. В., Костицына Н. В., Бакланов М. А. Влияние засоления и повышенной температуры воды на характеристики крови рыб в условиях бореальных водоемов: результаты эксперимента с использованием молоди речного окуня Perca fluviatilis бассейна р. Камы (Пермский край, Россия) // Вестник Астраханского государственного технического университета. Серия: Рыбное хозяйство. – 2022. – № 4. – С. 123–134.
- Шихмарев М. В. Влияние изменения температурного режима водоёма на жизнедеятельность водных экосистем // Актуальные научно-технические и экологические проблемы сохранения среды обитания: сборник материалов IV Международной научно-практической конференции (Брест, 25–27 сентября 2013 года) – Брест: БрГТУ, 2013. – С. 257–260.
- Ayllon F., Garcia-Vazquez E. Induction of micronuclei and other nuclear abnormalities in European minnow Phoxinus phoxinus and mollie Poecilia latipinna: an assessment of the fish micronucleus test // Mutation Research/Genetic Toxicology and Environmental Mutagenesis. – 2000. – Vol. 467, N 2. – P. 177–186.
- Hassan A. M., El Nahas A. F., Mahmoud S., Barakat M. E., Ammar A. Y. Thermal stress of ambient temperature modulate expression of stress and immune-related genes and DNA fragmentation in Nile tilapia (Oreochromis niloticus (Linnaeus, 1758)) // Applied Ecology & Environmental Research. – 2017. – Vol. 15, N 3. – 12 p.
- Arellano-García M. E., Torres-Bugarín O., García-García M. R., García-Flores D., Toledano-Magaña Y., Sanabria-Mora C. S., García-Ramos J. C. Genomic instability and cyto-genotoxic damage in animal species // IntechOpen. – 2021. – 19 p.
- Hayashi M. The micronucleus test – most widely used in vivo genotoxicity test. Review // Genes and Environment. – 2016. – Vol. 38. – 6 p.
- Raptis C. E., van Vliet M. T. H., Pfister S. Global thermal pollution of rivers from thermoelectric power plants // Environmental Research Letters. – 2016. – Vol. 11, N 10. – P. 104011.
- Sylvester J. R. Critical thermal maxima of three species of Hawaiian estuarine fish: a comparative study // Journal of Fish Biology. – 1975. – Vol. 7, N 2. – P. 257–262.
- Tang D., Chen M., Huang X., Zhang G., Zeng L., Zhang G., Wu S., Wang Y. SRplot: A free online platform for data visualization and graphing. PLoS One. 2023. – 18 (11). – e0294236.
- Verones F., Hanafiah M. M., Pfister S., Huijbregts M. A., Pelletier G. J., Koehler A. Characterization factors for thermal pollution in freshwater aquatic environments // Environmental Science & Technology. – 2010. – Vol. 44, N 24. – P. 9364–9369.