
ЦИТОМОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ПЫЛЬЦЫ ТИСА ЯГОДНОГО (TAXUS BACCATA L.) В УСЛОВИЯХ КУЛЬТУРЫ В ПРЕДГОРНОЙ ЗОНЕ КРЫМА
CYTOMORPHOLOGICAL CHARACTERISTICS OF POLLEN FROM COMMON YEW (TAXUS BACCATA L.) CULTIVATED IN FOOTHILL ZONE OF CRIMEA
JOURNAL: «EKOSISTEMY», Issue 39, 2024
Publication text (PDF): Download
UDK: 582.471(470.6)
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Zakharenko G. S.1, Alivapova S. S.2, Sevastyanov V. E.1, Salogub R. V.1
1Institute “Agrotechnological Academy” of the V.I. Vernadsky Crimean Federal University, Simferopol, Russia
2Botanical Garden named after N.V. Bagrov of the V.I. Vernadsky Crimean Federal University Simferopol, Russia
TYPE: Article
DOI: https://doi.org/10.29039/2413-1733-2024-39-63-72
PAGES: from 63 to 72
STATUS: Published
LANGUAGE: Russian
KEYWORDS: Taxus baccata, pollen, exine, intine, natural selection.
ABSTRACT (ENGLISH):
Normally developed pollen grains of the common yew (Taxus baccata L.) are a unicellular gametophyte in the pollen grain shell. According to the existing classification, they are characterized as a spheroidal, small-leptomed, radially symmetrical, sackless, rom small to medium size. Their diameters vary from 15.8 µ to 33.8 µ, with its average statistical value for individual trees from 21.1 µ to 27.2 µ. Cytomorphological study of pollen from the same yew trees in the Foothill zone conducted in 2015, 2016 and 2024 revealed that in timespans without significant temperature fluctuations during pollen formation and pollination, most yew trees produced pollen with an average viability of over 80 %. Conversely, in years with a sharp cold spells during the period when microstrobilus prepare for pollination or during pollination, most trees exhibited complete or partial damage to microstrobilus and a loss of their ability to disperse pollen. In some trees, that partially saved their ability to pollinate, pollen viability sharply decreases to 11–15 %. The most common occurred anomaly that reduces pollen viability is a disturbance of the structure of the pollen grain shell. As a result, pollen grains lose the ability to shed exine when they enter a pollination drop secreted by ovules during pollination. Non-shedding of exine excludes the participation of pollen with such anomaly in the sexual process, since during swelling the outer pectin layer of intine absorbs trophic and biologically active substances contained in the pollination drop, necessary for the growth of the pollen tube and the development of the male gametophyte. A decrease in the correlation between the average diameter of the pollen grain before the moment of shedding of the exine and their volume after shedding indicates the autonomization of the processes of formation of individual elements of the pollen grain shell. Significant differentiation of pollen grains by volume during swelling of the intine is considered a manifestation of natural selection in species with taxoid pollen. Data on the quality of pollen in yew and other gymnosperms, pollination of which is associated with the secretory activity of ovules, suggest that a minimum of three pollen grains is necessary for its normal pollination. It enhances the likelihood of producing viable seeds and facilitating the effects of natural selection in male sexual reproduction.
ВВЕДЕНИЕ
На территории России дизъюнктивный природный ареал тиса ягодного (Taxus baccata L.) охватывает горные районы Кавказа, Крыма и Калининградскую область, а в культуре этот вид широко распространён в теплоумеренных районах страны. В крымской части природного ареала и декоративных насаждениях полуострова этот, как правило, бипариентальный вид представлен деревьями обоего пола. Здесь как у женских, так и мужских у деревьев, вступивших в репродуктивную фазу, ежегодно закладывается большое количество репродуктивных органов. Однако, по нашим наблюдениям, в отдельные годы при совместном произрастании мужских и женских деревьев у тиса в декоративных насаждениях города Симферополя урожай семян отсутствовал.
Изучение репродуктивных способностей вида в условиях культуры является одним из критериев оценки его адаптивных возможностей и перспектив культивирования в новых физико-географических условиях (Нестерович, 1955; Мауринь, 1967). Семеношение в условиях культуры способствует повышению адаптивных возможностей древесных растений в череде семенных поколений и может обеспечить, как конечный результат акклиматизации, формирование интродукционных популяций, адекватных по своим экологическим требованиям условиям района массового выращивания (Мауринь, 1967; Некрасов, 1980).
Современные отечественные публикации, посвященные изучению биологии тиса ягодного, в основном касаются состояния насаждений, особенностей морфологии, экологии и естественного возобновления в горных лесах Осетии (Базаев, 2006), внутривидовой морфологической изменчивости и популяционной структуры этого вида в естественных насаждениях горного Дагестана (Омарова, Асадулаев, 2015; Омарова, 2018). В условиях интродукции в городе Махачкала средняя полнозернистость семян тиса составляет 94±4,25 % (Хазриева, 2011). В экспериментальных посевах установлено различие всхожести семян из природных популяций горных районов республики, варьирующей от 1,8 % до 16 % (Омарова, 2019). В природных насаждениях Западной Европы жизнеспособность семян тиса в среднем составляет 67 % с колебанием 4770 %. (Rudolf, 1974).
Изучение естественного возобновления и особенностей репродуктивного развития тиса в местах естественного произрастания в Крыму показало, что разновозрастность подроста и периодичность семеношения в локальных насаждениях в разных высотных зонах связана с нарушениями на разных этапах микроспорогенеза и снижением качества пыльцы, а также с опылением при неблагоприятных гидротермических условиях (Ругузов, 1978). В предгорной зоне Крыма, где тис ягодный по биологической сути является интродуцентом, при совпадении сроков поллинации с наступлением морозной погоды наблюдается гибель микростробилов, готовых к рассеиванию пыльцы, вследствие поражения гидратированных клеток их центральной оси и нераскрывания микроспорангиев (Захаренко, Зильберварг, 2018).
У тиса ягодного пыльца представляет собой микроспору в оболочке пыльцевого зерна. По морфологической классификации его пыльцевые зерна радиально симметричные мелколептомные, диаметром 22–25 мкм (Сладков, 1967). Экзина тонкая с мелкозернистой поверхностью. Интина толстая, состоит из наружного пектинового слоя и внутреннего слоя, содержащего целлюлозу (Müller-Stoll, 1948).
У тиса, как и у других голосеменных растений, у которых в рецептивной фазе семяпочки выделяют секреторную жидкость, нормальные пыльцевые зерна при попадании в опылительную каплю в результате гидратации пектинового слоя интины освобождаются от экзины. По этому признаку Мюллер-Штолль (Müller-Stoll, 1948) отнес пыльцу тиса и ряда других видов голосеменных растений к таксоидному типу.
Как показали исследования пыльцы и механизмов опыления у тиса ягодного (Никифоров, Ругузов, 1973) секвойи вечнозеленой (Никифоров Захаренко, 1973) и кипарисов (Захаренко, 2005), освобождение от экзины в опылительной жидкости, выделяемой семяпочками в рецептивной фазе, является необходимым этапом процесса опыления, последующего развития мужского гаметофита и оплодотворения. В связи с этим у видов голосеменных растений, механизм опыления которых связан с секреторной деятельностью семяпочек и сбрасыванием пыльцевыми зернами экзины, к нормально развитым пыльцевым зернам, способным участвовать в половом процессе, относятся лишь способные сбрасывать экзину в опылительной капле или имитирующей ее водной среде.
Большинство декоративных форм тиса ягодного, широко используемых в декоративном садоводстве, отобраны среди саженцев семенного происхождения в питомниках. В связи с тем, что тис ягодный относится к охраняемым растениям, возможности привлечения семян из мест природного обитания ограничены. По этой причине источником посевного материала для налаживания его размножения и введения селекционной работы служить без ограничений декоративные насаждения этого вида. При этом отметим, что семенное размножение растений обеспечивает микроэволюцию и формирование генофонда, коадаптированного физико-географическим условиям района интродукции.
В целях выяснения причин нерегулярного семеношения и оценки репродуктивных возможностей тиса ягодного в предгорном Крыму нами изучены морфофизиологические особенности его пыльцы, формирующейся в данном районе культуры.
МАТЕРИАЛ И МЕТОДЫ
Объектом исследования служили образцы пыльцы тиса ягодного, собранные в 2015, 2016 и 2024 годах с деревьев, растущих на территории Агротехнологической академии, Ботанического сада Крымского федерального университета. Для оценки способности пыльцевых зерен сбрасывать экзину пыльцу предварительно на 45 минут помещали в 0,1-молярный раствор уксуснокислого кальция, после чего раствор удаляли с помощью фильтровальной бумаги, а пыльцу окрашивали ацетокармином (Захаренко, Ругузов, 1982). На полученных препаратах под микроскопом МБР-1 с помощью окуляр-микрометра МОВ 1-1,5× с точностью до 0,1 микрона у пыльцевых зерен, сбросивших экзину, измеряли диаметр и не менее чем в пятнадцати полях зрения подсчитывали число пыльцевых зёрен, сбросивших и не сбросивших экзину, а также других аномальных пыльцевых зерен. Для определения нативных размеров летящей пыльцы при приготовлении окрашенных препаратов исключали её предварительную обработку физиологическим раствором.
Математическая обработка полученных результатов осуществлена с использование пакета прикладных программ Microsoft Excel 2003 и Statistica. Оценка уровня изменчивости признаков дана по шкале С. А. Мамаева (Мамаев, 1973).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
По данным наших наблюдений, поллинация у тиса ягодного в предгорном Крыму идет в позднезимний-ранневесенний период, в дни с положительными среднесуточными температурами +5–7 ºС и дневными повышениями до +10–11 °С. В 2015 году пыление у отдельных деревьев на территории парка Агротехнологической академии шло в первой декаде марта, а в 2024 году в середине третьей декады февраля. В пределах кроны отдельного дерева тиса рассеивание пыльцы длится около 5 суток. При этом отметим, что в данном районе культуры тису свойственна протогиния. Переход семяпочек в рецептивное состояние, характеризуемое началом секреторной деятельности семяпочек, наступил на 3–5 дней раньше начала поллинации.
Рассеивание пыльцы из микроспорангиев происходит после растяжения (роста) оси и расхождения микроспорофиллов микростробила и длится в пределах отдельного микростробила от нескольких минут в теплую сухую погоду до трех – шести часов при температуре +10–11 °С. При падении температуры до −2–3 °С у распускающихся микростробилов повреждаются растягивающиеся гидратированные клетки оси стробилов и теряется способность рассеивать пыльцу.
В 2015 году, в результате воздействия отрицательных температур, у большинства мужских деревьев тиса ягодного, растущих на территории Ботанического сада КФУ, несмотря на обильную закладку микростробилов, пыление не наблюдалось. В то же время у деревьев, растущих на территории ветеринарного факультета Агротехнологической академии, наблюдалось нормальное рассеивание пыльцы. У дерева же, растущего внутреннем дворе Агротехнологической академии между корпусами 1 и 2, способность пыления сохранила только часть микростробилов. Как показал анализ, в ночь на 3 марта 2015 года оказались пораженными около 45 % микростробилов, расположенных в базальной части побегов. В 2016 году температурный режим был благоприятен для поллинации всех деревьев тиса в рассматриваемом районе. В 2024 году на территории Агротехнологической академии погодно-климатические условия и ход поллинации тиса, проходившей в третьей декаде февраля, были аналогичны ситуации, наблюдаемой в 2015 году.
Зрелое пыльцевое зерно тиса ягодного имеет шаровидную форму и представляет собой микроспору в оболочке пыльцевого зерна. Ядро микроспоры занимает центральное положение и окружено структурированной цитоплазмой с мелкими вакуолями (рис. 1).
Цитоморфологическое изучение образцов зрелой пыльцы, результаты которого представлены в таблице 1, показало, что у тиса ягодного в условиях Предгорной зоны Крыма формируется морфологически нормальная пыльца. У большинства деревьев (у 9 из 11) доля пыльцевых зерен, сбрасывающих экзину и не имеющих видимых аномалий ядра и цитоплазмы, в годы исследования составляет от 69,7 % до 98,3 %. Лишь в образцах пыльцы двух деревьев (№ 1 и № 4), имевших в верхней части побегов микростробилы, не поврежденные морозом и не утратившие способность к поллинации, в 2015 году составила менее 15 %.
![]() а
а 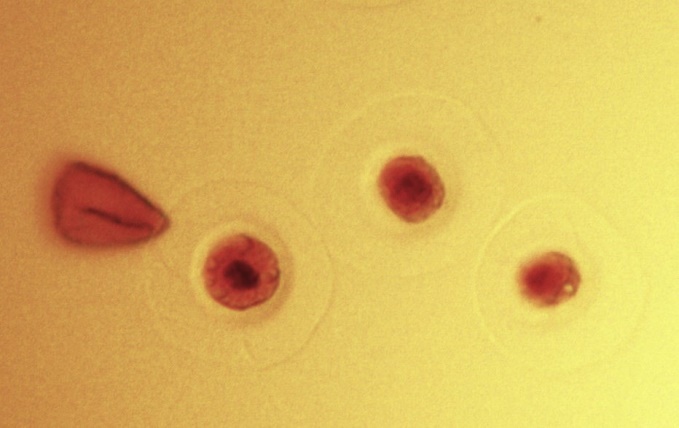 б
б
Рис. 1. Тис ягодный (Taxus baccata L.)
а – микростробилы на ветке дерева; б – пыльцевые зёрна тиса ягодного, сбросившие экзину, в левой части сброшенная экзина.
Использованная методика приготовления препаратов, имитирующая процесс опыления, выявила, что основной аномалией, снижающей качество пыльцы у этих деревьев, является неспособность части пыльцевых зерен, не имеющих видимых морфологических нарушений, сбрасывать экзину в физиологическом растворе, т.е. неспособность участвовать в половом процессе. В наиболее репрезентативной выборке пыльцы десяти деревьев в 2016 году количество пыльцевых зерен с такой аномалией варьировало от 1,1 % (дерево № 10) до 11,3 % (дерево № 2).
Наибольшее количество пыльцевых зерен с такой аномалией (от 17,7 % до 70,3 %) отмечено в 2015 году в образцах пыльцы деревьев (№ 1, № 3, № 4; № 6) с частично неповреждёнными микростробилами, когда у абсолютного большинства деревьев тиса вследствие морозного поражения микростробилов пыление отсутствовало. При этом отметим, что у в образце пыльцы дерева № 2, растущего в защищенном месте на территории Агротехнологической академии, в 2015 году количество пыльцы с этой аномалией составляло 3,1 %, что значительно ниже этого показателя (10,9 %) в 2016 году, благоприятном по климатическим условиям для поллинации.
Среди пыльцевых зерен, неспособных сбрасывать экзину, также встречались нераспавшиеся диады с живой цитоплазмой и ядрами (от единичного количества до 1,1 % у дерева № 1 в 2015 году), а также оптически пустые зерна (до 5,4 % у дерева № 10 в 2016 году).
В 2015 году в образцах пыльцы четырёх из пяти деревьев с частично повреждёнными микростробилами обнаружены нераспавшиеся тетрады, среди которых преобладали отмершие на ранних этапах развития (до 40,5 % у дерева № 1). У них наблюдалось отсутствие сформированных внешних оболочек и клеточного содержимого. Судя по неразвитости внешних слоёв оболочки, тетрады погибли вскоре после второго шага мейоза. Это наблюдалось у особей, растущих на как на территории Агротехнологической академии, так и Ботанического сада Крымского федерального университета.
Причины возникновения аномальных пыльцевых зерен разные. Можно полагать, что неспособность сбрасывания экзины связана как с формированием более толстой и прочной экзины, так и нарушениями развития внешнего слоя пектинового слоя интины, что может быть проверено лишь путём исследования строения оболочки пыльцевого зерна. Наличие нераспавшихся диад внешне нормальных микроспор связано с нарушениями в процессе формирования тетрад, идущем по сукцессионному типу. Более глубокие нарушения в процессе мейоза и формирования тетрад приводят к отмиранию микроспор непосредственно после завершения формирования их первичных клеточных оболочек. Поскольку у тиса ягодного в Крыму мейоз микроспороцитов идет в зимний период, когда возможны резкие понижения температур, часть аномалий развития пыльцы и отмирания пыльцевых зерен на разных этапах развития обусловлены неблагоприятными погодными условиями в период прохождения кинетических этапов мейоза. Присутствие в отдельных образцах оптически пустых пыльцевых зерен с развитой оболочкой, близких по размерам нормальным, по-видимому, также связано с нарушениями в мейозе, вызванными низкими температурами. Косвенным подтверждением этого являются данные, приводимые И. А. Ругузовым (1972), наблюдавшим у тиса ягодного в культуре на Южном берегу Крыма и в природных популяциях в Горном Крыму широкий спектр нарушений в процессе редукционного деления и формирования микроспор, в том числе гибель части из них на поздних этапах развития.
Таблица 1
Цитоморфологическая характеристика пыльцы у тиса ягодного в условиях культуры в городе Симферополь
| № дерева | Год | Сбрасывающая экзину, % | Неспособная сбрасывать экзину, % | Отмершая тетрада микроспор, % | |||||
| Морфологи-чески нормальная | Взрывающаяся | Отмершая | Морфологи-чески нормальная | Нераспав-шиеся диады | Оптически пустая | ||||
| На ранних этапах | На поздних этапах | ||||||||
| 1 | 2015 | 11,1 | ед. | 0,8 | 43,6 | 1,1 | 1,1 | 40,5 | 1,8 |
| 2016 | 89,5 | 0,4 | 4,8 | 4,6 | — | 0,7 | — | — | |
| 2024 | 85,1 | ед. | 4,0 | 10,9 | ед. | ед. | ед. | — | |
| 2 | 2015 | 87,5 | ед. | 1,0 | 3,1 | 0,5 | 0,7 | 7,2 | — |
| 2016 | 85,3 | ед. | 3,3 | 11,3 | — | 0,1 | — | — | |
| 2024 | 87,8 | ед. | 11,3 | 0,1 | ед. | 0,8 | — | ед. | |
| 3 | 2015 | 69,7 | ед. | 1,0 | 27,6 | 0,4 | 1,3 | — | — |
| 2016 | 79,8 | — | 9,8 | 9,8 | ед. | 0,6 | ед. | ед. | |
| 4 | 2015 | 14,9 | ед. | 0,8 | 70,3 | — | 5,2 | 7,9 | 0,9 |
| 2016 | 84,2 | — | 4,9 | 10,8 | ед. | 0,1 | — | — | |
| 5 | 2016 | 89,3 | 0,1 | 5,0 | 4,2 | — | 0,1 | 1,0 | 0,3 |
| 6 | 2015 | 78,3 | ед. | 2,2 | 17,7 | — | 1,1 | 0,7 | — |
| 7 | 2016 | 94,0 | ед. | 3,9 | 1,7 | — | 0,4 | — | — |
| 8 | 2016 | 92,7 | ед. | 2,6 | 4,7 | — | ед. | — | — |
| 9 | 2016 | 86,0 | ед. | 2,2 | 9,1 | — | 2,7 | — | — |
| 10 | 2016 | 91,4 | ед. | 2,1 | 1,1 | — | 5,4 | — | — |
| 11 | 2016 | 93,3 | ед. | 2,9 | 3,4 | 0,1 | 0,3 | — | — |
Кроме описанных выше аномалий, на препаратах пыльцы, обработанной физиологическим раствором, наблюдались единичные зерна, у которых при сбрасывании экзины разрывался внутренний слой интины. При этом цитоплазма часто вместе с ядром изливались в разбухший слой интины. Такие пыльцевые зерна мы относим к неспособным к формированию трубки и участию в половом процессе.
Результаты цитоморфологического анализа показали, что в условиях культуры в предгорной зоне Крыма у тиса ягодного у большинства деревьев формируется морфологически нормальная пыльца с высокой жизнеспособностью, превышающей 80 % (при колебании от 69,7 % до 94,0 %). Лишь в годы с экстремальными погодными условиями в период микроспорогенеза и поллинации, когда у большинства деревьев имеет место полное поражение микростробилов, у отдельных деревьев, вероятно, отличающихся по ритму развития микростробилов, при частичном поражении микростробилов жизнеспособность пыльцы падает до 11,1 % и 14,9 % у деревьев № 1 и № 4 или даже сохраняет достаточно высокий уровень жизнеспособности как у деревьев № 2, № 3 и № 6 в 2015 году и у дерева № 1 в 2024 году.
Данные о качестве пыльцы у одних и тех же деревьев в разные годы позволяют видеть, что в условиях нестабильного термического режима позднезимнего и ранневесеннего периода важным фактором, обеспечивающим возможность успешного опыления у тиса в предгорной зоне, являются как фенологическая гетерогенность деревьев, так и разновременность рассеивания пыльцы микростробилами отдельно взятого побега. Неодновременная поллинация микростробилов в пределах одного побега, идущая в акропетальной последовательности, расширяет возможность частичного сохранения жизнеспособности пыльцы и опыления даже при поражении низкими температурами части готовых к поллинации микростробилов.
Результаты измерения диаметра пыльцевых зёрен, не подверженных обработке физиологическим раствором (таблица 2), показывают, что в условиях Предгорной зоны Крыма средний диаметр пыльцевого зерна в изученных образцах варьирует от 21,1±0,27 мк до 27,2±0,32 мк при абсолютных значениях этого показателя от 15,8 мк (дерево № 4) до 33,8 мк (дерево № 6). Во всех изученных образцах пыльцевые зерна достаточно выравнены, по диаметру и по шкале С. А. Мамаева этот морфологический признак оценивается низким уровнем изменчивости (8≤С≤11%).
Сравнение средних, минимальных и максимальных значений диаметра пыльцевого зерна показывает, что наиболее крупные пыльцевые зерна превышаю по диаметру мелкие в среднем в 1,6 раза. Различия между отдельными зёрнами в пределах образца более явны при сравнении их объёмов: пыльцевые зерна с максимальным значением диаметра в среднем более чем в 4,3 раза превышают по объёму мелкие.
Дисперсионный анализ данных измерения диаметра пыльцевого зерна в наиболее репрезентативной выборке 2016 года выявил существенные различия деревьев рассматриваемой группы по этому показателю: Fфактическое =52,10 при Fкритическое=1,96 и Р<0,0001.
Результаты измерения диаметра и расчёта объёма пыльцевых зёрен после сбрасывания экзины в физиологическом растворе, приведенные в таблице 3, показывают, что в результате разбухания наружного слоя интины пыльцевые зерна увеличиваются в диаметре в 1,5–1,7 раза. Среднестатистическое значение диаметра зерна в рассматриваемых образцах варьирует от 32,7 мк (дерево № 2 в 2015 году) до 43,7 мк (дерево № 9 в 2016 году) при абсолютных значениях признака от 24,5 мк (дерево № 1 в 2024 году) до 57,8 мк (дерево № 4 в 2015 году). Варьирование пыльцевых зёрен, сбросивших экзину, по диаметру несколько возрастает и характеризуется уровнем изменчивости от низкого до среднего (9≤С≤20 %).
Объем пыльцевого зерна при сбрасывании экзины в среднем увеличивается в 3,6–5,6 раза. В изученных образцах пыльцы среднестатистическое значение этого признака колеблется от 19068 мк3 (дерево № 2) до 41167 мк3 (дерево № 7). Абсолютные значения объёма пыльцевого зерна, сбросившего экзину, изменяются в широких пределах и составляют от 9198 мк3 (деревья № 2 и № 8) до 100794 мк3. Если по среднему значению объёма пыльцевого зерна образцы отличаются в 2,2 раза, то по абсолютному значению этого показателя пыльцевые зерна в пределах рассмотренных образцов отличаются в 11 раз. У пыльцевых
Таблица 2
Средние значения диаметра и объёма пыльцевых зёрен с экзиной в образцах пыльцы деревьев тиса ягодного в условиях культуры в городе Симферополь
| № дерева | Год | Диаметр пыльцевого зерна, мк | Объем пыльцевого зерна, мк3 | ||||
| Dср.± m | Dмин-Dмакс. | С, % | V ср.± m | Vмин– Vмакс | С, % | ||
| 1 | 2015 | 24,5±0,31 | 19,0–29,5 | 10 | 7741,0±295,3 | 3589,5–13435,2 | 40 |
| 2016 | 24,9±0,28 | 16,3–29,9 | 11 | 8083,44±345,3 | 2226,1–13996,2 | 41 | |
| 2024 | 25,0±0,28 | 17,0–31,0 | 10 | 8181,2±0,329 | 2572,4–15598,4 | 38 | |
| 2 | 2015 | 22,4±0,30 | 18,5–26,5 | 11 | 6982,7±133,4 | 3315,2–9744,0 | 36 |
| 2016 | 22,9±0,28 | 17,4–28,0 | 11 | 6 287,9±176,1 | 2758,3–11494,0 | 40 | |
| 2024 | 23,1±0,29 | 17,3–27,9 | 9 | 6 454,1±189,3 | 2711,0–11371,3 | 34 | |
| 3 | 2015 | 23,0±0,25 | 19,2–27,8 | 9 | 6370,5±212,1 | 3733,1–11183,2 | 28 |
| 2016 | 24,0±0,26 | 20,0–27,7 | 10 | 7238,2±265,0 | 4188,8–11188,5 | 32 | |
| 4 | 2015 | 21,1±0,27 | 15,8–27,0 | 10 | 5044,3±194,2 | 2044,7–10300,8 | 50 |
| 2024 | 23,0±0,24 | 16,2–28,4 | 9 | 6370,6±224,2 | 6370,6–11993,7 | 44 | |
| 5 | 2016 | 24,8±0,27 | 19,5–30,0 | 9 | 8207,7±259,4 | 3880,5–14130,0 | 33 |
| 6 | 2015 | 27,0±0,32 | 20,8–33,8 | 10 | 10556,7±374,2 | 4676,6–20118,7 | 41 |
| 7 | 2016 | 26,0±0,31 | 20,8–32,2 | 10 | 9406,9±329,0 | 4675,6–17553,7 | 32 |
| 8 | 2016 | 24,2±0,23 | 20,0–29,2 | 8 | 7597,8±223,3 | 4186,7–13096,5 | 34 |
| 9 | 2016 | 27,2±0,32 | 22,0–32,8 | 10 | 10775,7±378,6 | 5572,4–18382,8 | 32 |
| 10 | 2016 | 25,2±0,26 | 16,2–33,2 | 11 | 8622,9±257,7 | 2192,2–19099,2 | 37 |
| 11 | 2016 | 23,7 ±0,22 | 17,1–29,2 | 9 | 7163,9±181,0 | 2593,9–13793,6 | 45 |
Таблица 3
Средние значения диаметра и объёма пыльцевых зёрен, сбросивших экзину, в образцах пыльцы деревьев тиса ягодного в городе Симферополь
| № дерева | Год | Диаметр пыльцевого зерна без экзины, мк | Объем пыльцевого зерна без экзины, мк3 | ||||
| Dср.± m | Dмин-Dмакс. | С, % | Vср.± m | Vмин–Vмакс | С, % | ||
| 1 | 2015 | 41,7±0,78 | 29,8–56,8 | 14 | 39862,1±2224,3 | 13779,7–95648,0 | 40 |
| 2016 | 40,2±0,63 | 27,6–53,4 | 13 | 34015,4±2021,5 | 11008,4–82417,4 | 36 | |
| 2024 | 40,4±0,52 | 24,5–55,9 | 14 | 35525,6±1918,9 | 7700,1–91460,4 | 38 | |
| 2 | 2015 | 32,7±0,46 | 26,0–45,5 | 11 | 19068,3±859,4 | 9198,1–49296,1 | 36 |
| 2016 | 35,8±0,69 | 25,3–54,8 | 9 | 24024,1±763,4 | 8479,3–86166,7 | 38 | |
| 2024 | 33,9±0,59 | 24,8–52,8 | 10 | 20398,4±799,5 | 7986,4–77072,5 | 35 | |
| 3 | 2015 | 39,9±0,45 | 29,25–48,5 | 9 | 34193,3±1129,9 | 13096,5–59704,0 | 28 |
| 2016 | 38,2±0,45 | 27,0–50,1 | 11 | 29186,9±1128,1 | 10306,0–65843,2 | 31 | |
| 4 | 2015 | 36,4±0,66 | 26,5–57,8 | 14 | 26746,7±1717,9 | 9739,0–100793,9 | 50 |
| 2016 | 37,6±0,58 | 26,1–46,8 | 13 | 27833,2±1418,3 | 9630,4–53670,4 | 41 | |
| 5 | 2016 | 39,6±0,53 | 30,5–50,3 | 11 | 33718,7±1361,1 | 14848,3–66402,8 | 33 |
| 6 | 2015 | 40,9±0,70 | 27,0–54,3 | 14 | 37880,9±1913,3 | 10300,7–83556,0 | 41 |
| 7 | 2016 | 42,3±0,63 | 24,8–52,8 | 9 | 41167,4±723,5 | 7934,2–76815,0 | 29 |
| 8 | 2016 | 37,3±0,54 | 26,0–47,0 | 13 | 28343,5±1188,3 | 9198,1–54334,0 | 34 |
| 9 | 2016 | 43,7±0,56 | 31,0–56,2 | 11 | 45137,6±1723,3 | 15590–93142,1 | 32 |
| 10 | 2016 | 39,9±0,64 | 30,7–50,2 | 13 | 34931,7±1594,4 | 15127,6–66284,0 | 37 |
| 11 | 2016 | 40,4.±0,88 | 28,5–56,5 | 15 | 36577,2±24722,4 | 12114,7– 94389,5 | 45 |
зёрен, сбросивших экзину, существенно возрастает и уровень варьирования этого признака – от высокого до очень высокого (29≤С≤50 %).
Сравнение результатов измерения диаметра и объёма пыльцевых зёрен до момента сбрасывания экзины и после освобождения от экзины в физиологическом растворе выявило достоверно выраженную корреляцию между этими показателями (табл. 4).
Таблица 4.
Значения коэффициента корреляции (r) между средними значениями диаметров и объёмов пыльцевых зёрен в пыльце деревьев тиса ягодного в городе Симферополь
| Показатели | Диаметра пыльцевого зерна с экзиной | Диаметр пыльцевого зерна без экзины | Объём пыльцевого зерна с экзиной |
| Диаметр пыльцевого зерна без экзины | 0,773 | ||
| Объём пыльцевого зерна с экзиной | 0,979 | 0,707 | |
| Объём пыльцевого зерна без экзины | 0,783 | 0,992 | 0,737 |
Наблюдается практически полная корреляция между средними значениями диаметров и объёмов пыльцевых зёрен как в образцах с экзиной, так и после сбрасывания экзины: значение коэффициента корреляции близко к единице (r≥0,979). Заметно ниже зависимость между диаметрами и объёмами пыльцевых зёрен до и после сбрасывания экзины (r≤0,8), а также между средними значениями объёма пыльцевых зёрен до и после сбрасывания экзины (r=0,737). Менее тесная корреляция между этими показателями, вероятно, указывает на различие пыльцевых зёрен по степени развития их наружного, разбухающего в опылительной капле или физиологическом растворе пектинового слоя интины при близких значениях их диаметров до сбрасывания экзины.
Как показал анализ химического состава секрета опылительной капли (“pollination drop”) в англоязычной литературе), выделяемого в рецептивной фазе семяпочками тиса, он содержит пластические и биологически активные вещества (Циглер, 1959). В результате сбрасывания экзины разбухающий слой интины является приспособлением для депонирования этих веществ, обеспечивающих дальнейшее формирование мужского гаметофита и не обладающей гаусториальными свойствами пыльцевой трубки, растущей между стенкой нуцеллуса и внутренней поверхностью интегумента. В этой связи дифференциацию по объёму пыльцевых зерен, сбрасывающих экзину, можно рассматривать как один из механизмов отбора в гаплофазе мужского гаметофита.
Сопоставление данных цитоморфологического анализа пыльцы голосеменных растений, опыление у которых связано с секреторной деятельностью семяпочек, показывает наличие одинаковых аномалий в зрелой пыльце. У тиса, как и у таксодиевых и кипарисовых, к числу основных аномалий, исключающих участие части пыльцевых зёрен в половом процессе, является их неспособность сбрасывать экзину. Наличие разного количества пыльцевых зёрен с такой аномалией, вероятно связано не столько с отрицательным воздействием низких температур на этапе подготовки к поллинации, как это проявилось 2015 году, но и с индивидуальными особенностями дерева и отдельных микроспор, имеющих определённую автономию развития, хотя и под контролем материнского дерева через физиолого-биохимическую среду, поддерживаемую в микроспорангии. Эта аномалия может быть обусловлена как избыточной прочностью экзины, так и недоразвитием пектинового слоя интины, что может быть установлено только при изучении строения и структуры оболочки пыльцевого зерна.
Данные о жизнеспособности пыльцы у тиса и других хвойных с аналогичным способом опыления позволяют объяснить тот факт, что для нормального опыления семяпочки требуется попадание в опылительную каплю минимально трёх пыльцевых зёрен, после чего секреторная деятельность семяпочки прекращается. Необходимо учитывать, что на этапе поллинации во всех изучаемых образцах пыльцы зрелой пыльцы невозможно точно определить реальную оплодотворяющую способность пыльцевых зёрен, классифицируемых как морфологически нормальные и жизнеспособные, поскольку не представляется возможным достоверно прогнозировать их последующее развитие до образования спермиев и оплодотворения. Наличие во всех образцах без исключения только части цитоморфологически нормальных пыльцевых зёрен, фертильность которых лишь частично достоверна на первых этапах развития мужского гаметофита, а также их очень высокая изменчивость по размерам разбухающего в опылительной капле пектинового слоя интины, важного для последующего питания и развития мужского гаметофита, позволяют рассматривать необходимость для нормального опыления семяпочки минимально трёх пыльцевых зёрен как страховку, расширяющую возможность не только производства жизнеспособных семян, но и действие естественного отбора в мужском половом поколении.
ЗАКЛЮЧЕНИЕ
У тиса ягодного в условиях культуры в Предгорной зоне Крыма в годы с благоприятными условиями прохождения всех этапов микроспорогенеза формируется нормально развитая одноклеточная пыльца с жизнеспособностью, превышающей 80 % и колебанием от 69,7 % до 94,0 %. В годы с экстремальными погодными условиями в период микроспорогенеза и поллинации наблюдается полное поражение микростробилов низкими температурами, но у отдельных деревьев, отличающихся по ритму развития, при частичном поражении микростробилов жизнеспособность пыльцы падает до 11,1–14,9 %.
У тиса ягодного в зрелой пыльце наблюдается весь спектр аномалий, характерных для видов кипарисовых и таксодиевых, у которых опыление также связано с секреторной деятельностью семяпочек в рецептивной фазе их развития. Основными аномалиями пыльцы являются: несбрасывание экзины при попадании в физиологический раствор, моделирующий секрет семяпочки, наличие отмерших микроспор на разных этапах развития от формирования тетрад до поллинации.
Качество пыльцы в годы с отсутствием заморозков в период поллинации потенциально обеспечивает нормальное семеношение тиса ягодного в декоративных насаждениях Предгорной зоны Крыма.
- Базаев А. Б. Тис ягодный в горных лесах Осетии: Особенности строения и возобновительный потенциал: Автореферат диссертации на соискание учёной степени кандидата биологических наук: 06.03.03. С-Пб, 2006,
18 с. - Захаренко Г. С. Особенности развития пыльцы секвойи вечнозеленой // Труды Никитского ботанического сада. – 1984. – Т. 92. – С. 84–91.
- Захаренко Г. С., Зильберварг И. Р. Фенологические особенности развития репродуктивных органов и завязываемость семян у тиса ягодного в предгорной зоне Крыма // Экология и география растений и растительных сообществ: материалы IV Международной научной конференции (Екатеринбург, 16–19 апреля 2018 года.). – Екатеринбург: Изд-во Урал. ун-та; Гуманитарный ун-т, 2018. – C. 309–311.
- Захаренко Г. С., Ругузов И. А. Особенности развития мужского гаметофита у Таксодиевых, Кипарисовых и Тисовых // Материалы Всесоюзного совещания «итолого-эмбриологические и генетико-биологические основы опыления и оплодотворения растений». – Киев: Наукова думка, 1982. – С. 222–225.
- Мамаев С. А. Формы внутривидовой изменчивости древесных растений (на примере семейства Pinaceae на Урале). – М.: Наука, 1973. – 284 с.
- Мауринь А. М. Семеношение древесных экзотов в Латвийской ССР. – Рига: Звайгзне, 1967. – 208 с.
- Некрасов В. И. Основы семеноведения древесных растений при интродукции. – М.: Наука, 1973. – 279 с.
- Некрасов В. И. Актуальные вопросы развития теории акклиматизации растений. – М.: Наука, 1980. – 102 с.
- Нестерович Н. Д. Плодоношение интродуцированных древесных растений в БССР. – Минск: Изд. АН БССР, 1955. – 382 с.
- Никифоров Ю. Л., Захаренко Г. С. Опыление у секвойи вечнозеленой // Половая репродукция хвойных. – Новосибирск: Наука. – 1973. – С. 139–141.
- Никифоров Ю. Л., Ругузов И.А. Морфогенез генеративных органов и опыление у тиса ягодного // Половая репродукция хвойных. – Новосибирск: Наука. – 1973. – С. 135–138.
- Омарова П. К. Изменчивость признаков шишкоягод Taxus baccata L. во внутреннегорном Дагестане // Ботанический вестник Северного Кавказа. – 2018. – № 1. – С. 20–25.
- Омарова П. К. Результаты размножения Тaxus baccata L. в Дагестане // Ботанический вестник Северного Кавказа. – 2019. – № 4. – С. 79–84.
- Омарова П. К., Асадулаев З. М. Изменчивость признаков шишкоягод Taxus baccata в популяциях Дагестана // Научный журнал КубГАУ, – 2015. – № 112 (08), 17 с.
- Ругузов И. А. Фертильность пыльцы тиса ягодного и аномалии в ее развитии // Бюллетень Государственного Никитского ботанического сада. – № 2 (18). – 1972. – С. 34–36.
- Ругузов И. А. Некоторые популяционно-морфологические аспекты сохранения тиса ягодного в Крыму // 50 лет Черноморскому государственному заповеднику. Материалы республиканского семинара-совещания. – Киев: Наукова думка, 1978. – С. 134–136.
- Сладков А. Н. Введение в споро-пыльцевой анализ. – М.: Наука, 1967, 269 с.
- Хазриева М. Р. Семенная продуктивность интродуцированных Pinopsida в условиях Махачкалы // Известия Дагестанского государственного педагогического университета. Естественные и точные науки. – 2011. – №2 (15). – С. 30–34.
- Müller-Stoll R. W. Zitomorphologische studien am Pollen von Taxus baccata L. und anderen Koniferen // Planta. –1948. – Bd. 35. – Hf. 5/6. – S. 601–641.
- Rudolf P. O. Taxus. Seeds of Woody Plants in the United States // Agriculture Handbook. – 1974. – N 450. – Р. 799–802.
- Ziegler H. Über die Zusammensetzung des «Bestaubungstropfens» und den Mechanismus seines Sekretion // Planta. – 1959. – Bd. 52. – S. 587–599.