
|
Экосистемы, 36: 234–247 (2023) |
О журнале |
|
УДК 581.93:574.3(470.6-924.71) |
DOI 10.5281/zenodo.10389713 |
Структурно-ценотические особенности ценопопуляций и анатомическое строение вегетативных органов Paronуchia cephalotes (M. Bieb.) Besser
Вахрушева Л. П., Николенко В. В., Петришина Н. Н., Котов С. Ф.
Крымский федеральный университет имени В. И. Вернадского Симферополь, Россия
vakhl@inbox.ru; niki-vera@mail.ru; n-petrishina@list.ru; sfktv@mail.ru
- статье представлены результаты изучения 6 ценопопуляций Paronуchia cephalotes (M. Bieb.) Besser, произрастающих в петрофитных степях и на петрофитоне известняковых скал предгорной части Крыма. При проективном покрытии 7 % и более P. cephalotes являлась содоминантом, численность ценопопуляций соответствовала 17–98 экземплярам, плотность 3,4 1,2 на 1 м2; при покрытии 1,0 % вид выполнял роль ассектатора, плотность составляла 0,9 0,6 особей на 1 м2. Все ЦП имели контагиозный тип пространственной структуры, образуя мономодальные (пик на генеративных особях) преимущественно неполночленные спектры. По классификации «дельта-омега» ценопопуляции относились к молодым (ЦП 5, 6) и находились на границе молодые-переходные (ЦП 1, 4), и типичным, переходным к зрелым (ЦП 2, 3). Некоторые данные об анатомическом строении вегетативных органов P. cephalotes получены впервые. Эти данные являются важным дополнением к общетеоретическим сведениям о внутриструктурной организации родов семейства Cariophyllaceae и позволили доказать принадлежность изученного вида к экологической группе ксеромезофитов и биоморфе полукустарничков.
Ключевые слова: Paronуchia cephalotes, ценопопуляция, охраняемый вид, пространственная структура, онтогенетическая структура, анатомическая структура, Крым.
ВВЕДЕНИЕ
Для Крыма, как одного из центров биоразнообразия, имеет особую актуальность сохранение редких видов растений в их природных популяциях в среде естественного произрастания (Meffe et al., 1994; Выработка приоритетов…, 1999; Примак, 2002). В рамках реализации Стратегического плана Международной Конвенции Декады Биоразнообразия, 2011–2020 гг. (Новости ООН, 2023; Convention on Biologycal Diversity, 2023) крымскими учеными накоплен значительный материал о структуре популяций редких видов флоры, необходимый для их охраны in situ (Брынза, 2014; Вахрушева, Абдулганиева, 2015; Летухова, Потапенко, 2017; Вахрушева, Заднепровская, 1918; Абдулганиева, 2020; Вахрушева, Абдулганиева, 2020; Vakhrusheva, Abdulganieva, 2020).
Комплексную морфолого-онтогенетическую оценку редких растений в последние годы стали обогащать и анатомическими исследованиями (Вахрушева, Николенко, 2017; Абдулганиева, 2020; Вахрушева, Петришина, Павшенко, 2020). В таком же комплексном формате было проведено изучение приноготовника головчатого (Paronуchia cephalotes (M. Bieb.) Besser) – вида, внесенного в Красную книгу Республики Крым (2016), Красную книгу города Севастополя (2018), Красную книгу Краснодарского края (2017) и Красную книгу Российской Федерации (2008), где для него в целом указываются низкая численность и плотность, но отмечается отсутствие точных данных об этих показателях, что свидетельствует о малой изученности популяций. Усилившееся в последние годы уничтожение мест обитаний приноготовника головчатого, в связи с интенсификацией хозяйственного освоения предгорий и строительством автомобильных трасс, обусловливает особую важность точных сведений о современном состоянии популяций P. cephalotes. Оказалась недостаточно исследованной и анатомическая структура не только
ISSN 2414-4738 Published by V. I. Vernadsky Crimean Federal University, Simferopol
Структурно-ценотические особенности ценопопуляций и анатомическое строение
вегетативных органов Paronуchia cephalotes (M. Bieb.) Besser
рассматриваемого вида, но и рода в целом (Borhidi, 1966; Schweingruber, 2007; Schweingruber et al., 2011; Schweingruber, Börner, 2018).
Цель настоящей работы – выяснить особенности пространственной и онтогенетической структур крымской популяции приноготовника головчатого (P. cephalotes), определить численность и плотность в разных эколого-ценотических условиях, изучить анатомическое строение вегетативных органов растений, оценить современное состояние его крымских ценопопуляций.
МАТЕРИАЛ И МЕТОДЫ
P. cephalotes – стелющийся стержнекорневой рыхлодерновинный полукустарничек высотой 5–10 см. Это понтический элемент флоры, аэропедофит, литофит, мезоксерофит и гелиофит (Голубев, 1996). В Крыму произрастает на щебнистых экотопах и поверхности скал степной и горной частях, общий ареал захватывает Балканы, Юго-Восточную Европу, Малую Азию, Западное Закавказье (окрестности Новороссийска). Природоохранный статус в Красной книге Российской Федерации (2008) указывается как вид, «сокращающийся в численности». Для Крымского полуострова отмечается как «вид вне опасности» (Красная книга Республики Крым…, 2016) и вид, «сокращающийся в численности» (Красная книга города Севастополя, 2018).
Изучение фитоценозов с произрастанием P. cephalotes проводились в 2018, 2019 и 2020 годы по стандартным геоботаническим методам: определялись флористический состав, проективное покрытие видов, устанавливалась синтаксономическая принадлежность сообществ, численность и плотность P. cephalotes (Шенников, 1964; Уранов, 1975). Пространственная структура анализировалась по значениям индекса Одума (Io) (Одум, 1986). Для установления онтогенетической структуры предварительно были выявлены критерии возрастных состояний P. cephalotes по Т. А. Работнову (1950), Л. А. Жуковой (2001). Онтогенетическая характеристика ценопопуляций в разных эколого-ценотических условиях оценивалась по комплексу индексов (Iв , Iс , Iг) (Коваленко, 2005). Возрастность (Δ – индекс возрастности) и эффективная плотность (ω – индекс эффективности) ценопопуляций определялись по методикам А. А. Уранова (1975) и Л. А. Животовского (2001). Совместное использование этих индексов было положено в основу установления типа ценопопуляций: молодые, зреющие, зрелые, переходные, стареющие и старые.
Анатомо-морфологические структуры надземной части растений изучались на фиксированном (спирт : глицерин : вода в соотношении 1 : 1 : 1) и нативном материале, собранном в фазе цветения. Анатомическое изучение проводили по общепринятым методикам (Прозина, 1960; Барыкина, 2004) на препаратах, полученных с помощью микротома Ротмик 2-П. Данные обрабатывались по стандартным методам математической статистики (Лакин, 1980). Исследование временных и постоянных препаратов проведены с использованием микроскопа Olympus CX31RTSF. Объекты фотографировали цифровой камерой Olympus (Industrial Digital Camera TOUPCAMTM U3CMOS10000KPA).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Фитоценозы, в составе которых произрастают изученные ценопопуляции P. cephalotes, приурочены к щебнисто-скальным экотопам предгорного Крыма. Большая часть ценопопуляций встречается в петрофитных степях и фриганоидных сообществах, однако иногда особи изучаемого вида выходят на скалы и известняковые глыбы, располагающиеся в пределах указанных фитоценозов, и формируют там самостоятельные ценопопуляции, достаточно четко отличающиеся по структуре от основного фона растительности. Подчеркнем – вся совокупность особей в этом случае рассматривается нами, как единая популяция (Seybold, 1980). Общее количество особей на момент исследования в пределах изученных ценопопуляций колебалось от 17 до 98 экземпляров, среднее число на 1 м2 зависело от роли вида в формируемом сообществе: если количественное участие P. cephalotes
- фитоценозе достигало значения содоминанта, то плотность составляла 3,4 1,1 на 1 м2, (проективное покрытие вида достигает при этом около 7 % и более); когда вид выполнял лишь роль ассектатора, то его плотность едва достигала 0,9 0,6 особей на 1 м2, а проективное покрытие было крайне незначительным – около 1,0 % (табл. 1).
Таблица 1
Геоботанические показатели фитоценозов и ценопопуляций Paronychia cephalotes
|
Ценопопуляция |
Площадь, м2 |
Фито |
||||
|
Тип экотопа / |
Название |
Численность / |
ценотическая |
|||
|
Покрытие |
плотность |
роль вида в |
||||
|
местоположение |
ассоциации |
|||||
|
травостоя / |
на1м2 |
составе данной |
||||
|
вида, % |
ассоциации |
|||||
|
Петрофитная степь |
||||||
|
(окрестности |
Bothriochloetum |
100 |
||||
|
г. Симферополь, |
Fumaneto- |
|||||
|
ЦП 1 |
44/0,3 |
ассектатор |
||||
|
ландшафтно- |
onosmeto- |
|||||
|
64,0/0,6 |
||||||
|
рекреационный парк |
genistosum |
|||||
|
«Битак») |
||||||
|
Известняковая скала |
||||||
|
(окрестности |
15 |
|||||
|
г. Симферополь, |
Thymoso- |
|||||
|
ЦП 2 |
17/0,9 |
cодоминант |
||||
|
ландшафтно- |
paronychiosum |
|||||
|
30,0/3,5 |
||||||
|
рекреационный парк |
||||||
|
«Битак») |
||||||
|
Петрофитная степь |
Festucetum- |
80 |
98,0/4,0 |
|||
|
(Симферопольский |
(в 2-х локусах), |
|||||
|
ЦП 3 |
Saturеetо |
содоминант |
||||
|
район, окрестности |
везде – |
|||||
|
paronychiosum |
60,0/7,0 |
|||||
|
с. Каменка) |
единично |
|||||
|
Петрофитная степь |
Festucetum |
100 |
||||
|
(Белогорский районн, |
Satureeto– |
|||||
|
ЦП 4 |
86,0/0,6 |
ассектатор |
||||
|
окрестности пещеры |
potentillietо- |
|||||
|
64,0/0,4 |
||||||
|
«Таврида») |
thymosum |
|||||
|
Известняковая скала |
Saturеetum |
9 |
||||
|
(Белогорский район, |
содоминант |
|||||
|
ЦП 5 |
paronychiosо- |
27,0/3,0 |
||||
|
окрестности |
(ассектатор) |
|||||
|
pleuroziosum |
80,0/3,4 |
|||||
|
с. Синекаменка) |
||||||
|
Петрофитная степь |
100 |
|||||
|
(Белогорский район, |
Stipetum-Saturеetо |
|||||
|
ЦП 6 |
88,0/1,4 |
содоминант |
||||
|
окрестности |
paronychiosum |
|||||
|
68,0/8,2 |
||||||
|
с. Синекаменка) |
||||||
Структура фитоценозов с участием P. cephalotes на известняковых скалах, несмотря на общность механического и химического состава экотопов, принципиально отличается по комплексу эколого-ценотических условий. Известняковая скала в парке «Битак» (ЦП 2) располагается на склоне юго-юго-западной экспозиции, основную часть светового дня она находится под прямыми лучами солнечного света, растения на ней произрастают лишь в небольших углублениях и кавернах известняка, поверхность только на 30 % покрыта растениями. Доминирует Thymus rogneri K. Koch (11,0 % проективного покрытия), покрытие P. cephalotes в этом фрагменте достигает до 3,5 %, он имеет невысокую численность и плотность (табл. 1). Известняковая скала в окрестности села Синекаменка (ЦП 5) находится также в пределах петрофитной степи, но весь участок располагается на пологом склоне северной экспозиции, а петрофитная степь по периметру окружена лесным массивом. Непосредственно поверхность самой скалы находится большую часть светового дня в густой тени, а проективное покрытие в этой ассоциации определяется не цветковыми растениями, а мхом Рleurozium schreberi (Brid.) Mitt. Отмеченные нами в ассоциации Satureja taurica Velen.
- P. cephalotes играют, по сравнению с мхом, подчиненную роль, достигая каждый покрытия около 3,4–4,6 % из 80,0 % общего проективного покрытия, то есть строго по фитоценотической роли в этом сообществе – это ассектаторы. Это тем более подчеркивает и эдификаторную роль мха в формировании биотопа – он накапливает влагу, долго сохраняет ее даже в самые засушливое время, что снимает экстремальность влияния этого важного экологического фактора на ценопопуляцию P. cephalotes в период засухи.
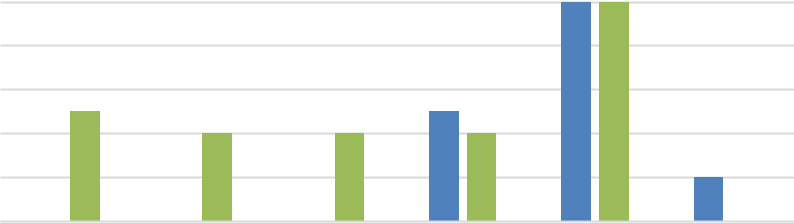
Отмеченные отличия в характере эко- и биотопов известняковых скал проявляются и в характере онтогенетической структуры произрастающих здесь ценопопуляций растений. Для исследования онтогенетической структуры нами был проведен морфометрический анализ признаков P. cephalotes, информативных для дифференциации его возрастных состояний. Выявлены следующие признаки: количество листьев, степень ветвления надземных побегов, смена типов нарастания побегов (моноподиального на симподиальное), характер их расположения в пространстве, соотношение вегетирующих и усохших побегов. На известняковой скале в парке «Битак» онтогенетический спектр (ЦП 2) неполночленный, мономодальный, с пиком на особях генеративного возрастного состояния. Левосторонняя часть его не содержит проростков, ювенильных и имматурных особей. Таким образом, имеет место депрессивная тенденция в развитии ЦП 2 (рис. 1).
На известняковой скале в окрестности села Синекаменка в ЦП 5 отсутствуют только особи сенильного возраста, вся левостороння часть полночленная, с достаточным количеством проростков (18,5 %), одинаковым участием других групп растений прегенеративного возраста (j, im, v – по 26,2 %), мономодальность спектра определяется четко выраженным преобладанием генеративных особей (рис. 1). Правостороння часть спектра характеризуется полным отсутствием на момент исследования субсенильных и сенильных растений, что и обусловливает его неполночленность (рис. 2). Статистически подтверждены на 95 % уровне значимости величины индексов Одума (Io): 1,18 (ЦП 2) и 1,21 (ЦП 5),
|
Количество, шт |
20
18
16
14
12
10
8
6
4
2
0
p j im v g s
Возрастное состояние
— Битак (известняковая глыба), ЦП 2
— Синекаменка (известняковая глыба), ЦП 5
Рис. 1. Онтогенетические спектры ценопопуляций Paronychia cephalotes (ЦП 2 и ЦП 5) свидетельствующие о групповом (контагиозном) характере произрастания особей в обеих ценопопуляциях.
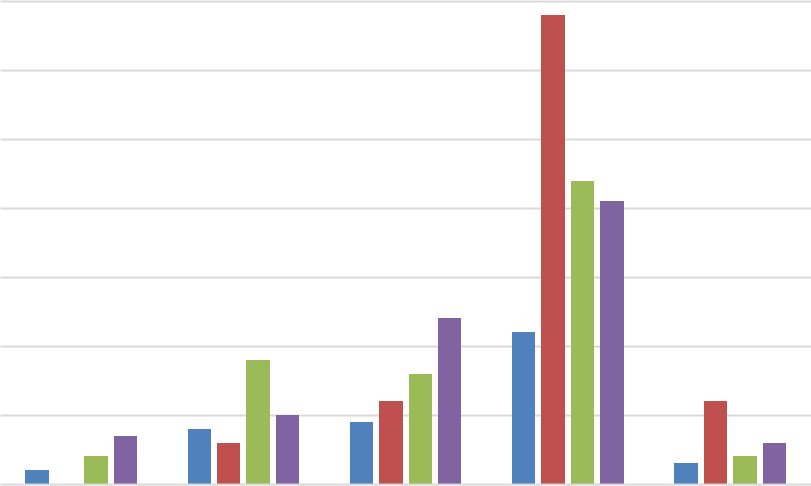
Онтогенетическая структура P. cephalotes во всех изученных фитоценозах петрофитных степей показывает следующие тенденции: очень мало особей в стадии проростков и ювенильном возрасте; с разными вариациями, но достаточное их количество в имматурном, виргинильном и сенильном возрастных состояниях и четко проявляется мономодальность всех спектров петрофитных степей с пиком на генеративных особях (рис. 2).
Из 6 изученных ценопопуляций только в петрофитной степи ЦП1 (рис. 2) и в петрофитоне известняковой скалы ЦП 5 (рис. 1) были найдены проростки; в трех – ЦП 4, 5, 6 были обнаружены ювенильные растения, то есть особи самой молодой части ценопопуляции есть и в наскальном комплексе и в петрофитных степях. Закономерности динамики онтогенетической структуры наиболее четко удалось проследить на петрофитоне известняковой скалы в Синекаменке (ЦП 5): проростки и ювенильные растения успешно появляются внутри «подушек» мха, здесь взрослеют до виргинильного возраста, а с момента подключения вегетативного размножения, за счет разрастания выходят за пределы моховых локусов и занимают образовавшимися раметами близлежащие пространства. На экотопах, где отсутствуют дерновины мха, прорастание семян происходит и в петрофитных степях, и во фригане, и в наскальных сухих комплексах в кавернах и каррах, где имеют место скопление мелких остатков мелкозема (3–5 мм в диаметре) с небольшим количеством почвы, поскольку именно здесь и были обнаружены единичные ювенильные растения (ЦП 1, 4, 6). Дальнейшее освоение ими пространства происходит по такому же типу, как и на скалах: экспансия рамет на близлежащие участки и дальнейшее распространение по всей площади, пригодной для их произрастания. У P. cephalotes в каждом из изученных экобиотопов имеют место тенденции, сходные с ценопопуляцими Satureja montana ssp. taurica (Velen.) P. W. Ball (S. taurica Velen.). Особи самых молодых возрастных состояний остаются наиболее уязвимыми и эллиминируются в первую очередь как на петрофитоне скал (ЦП 2), так и в условиях
|
Количество, шт |
70
60
50
40
30
20
10
0
j im v g s
Возрастное состояние
— Битак (петрофитная степь), ЦП 1
— Каменка (петрофитная степь/плоский известняк), ЦП 3
— Таврида (петрофитная степь), ЦП 4
— Синекаменка (петрофитная степь), ЦП 6
Рис. 2. Онтогенетические спектры ценопопуляций Paronychia cephalotes (ЦП 1, 3, 4, 6) петрофитных степей (ЦП 3). По мере взросления особей ценопопуляции меняют k-стратегию растений прегенеративного периода на r-стратегию генеративного возраста, поэтому при появлении процессов старения особи теряют устойчивость к неблагоприятным факторам и быстро отмирают (Абдулганиева, Вахрушева, 2020). В отличие от сходного по экобиоморфе кальцепетрофита S. montana, особи сенильного возраста P. cephalotes присутствуют в небольшом количестве в пяти онтогенетических спектрах из шести ценопопуляций (кроме ЦП 5), что может свидетельствовать о большей устойчивости этого вида к произрастанию на экстремальных местообитаниях. Однако ЦП 5 является самой молодой и, возможно, ее онтогенез в данном сообществе еще не достиг той фазы популяционной волны, которая характеризуется появлением сенильных растений (индекс возрастности Δ, по А. А. Уранову, самый низкий и равен 0,21).
Расчет разнообразных онтогенетических индексов (Iв – индекс возобновляемости; Iс – индекс старения; Iг – индекс генеративности; Δ – индекс возрастности; ω – индекс эффективности), позволяет охарактеризовать общее состояние изученных ЦП, а их значения подтверждают объективность различий ценопопуляций одного вида в разных эколого-ценотических условиях (табл. 2).
Значение индекса возобновляемости является важнейшим для прогноза ее существования в будущем. В ценопопуляциях петрофитных степей (ЦП 1, 4, 6) Iв имеет среднее значение (41,5–46,6 %), однако для ЦП 3, из петрофитной степи, Iв показывает минимальное значение 18,4, что обусловлено ее произрастанием в условиях наиболее высокого антропогенного пресса (Котов, Вахрушева, Епихин, 2016). В целом экотоп известняковых скал не является оптимальным местообитанием для осуществления процессов возобновления вида, о чем свидетельствует значение этого индекса для ЦП 2 (Iв=29,4 %). Однако, если складываются уникальные экологические условия, подобные скальному экотопу в окрестностях села Синекаменка (ЦП 5), тогда процессы возобновляемости протекают наиболее успешно и этот индекс достигает в данной выборке наибольшего значения (Iв=63,0). Индексы старения (Iс) показывают те же закономерности для изученных ЦП, но имеют значения противоположной направленности: если высок Iв, то низким будет Iс и наоборот (табл. 2).
ЦП 3, находящаяся в зоне сильнейшего антропогенного воздействия, имеет самый высокий индекс генеративности (Iг=69,4), который подтверждает наибольшую устойчивость этой возрастной группы для выживания в самых экстремальных условиях, поскольку устойчивость малых популяций редких видов определяется именно значениями коэффициента генеративности (Злобин и др., 2013).
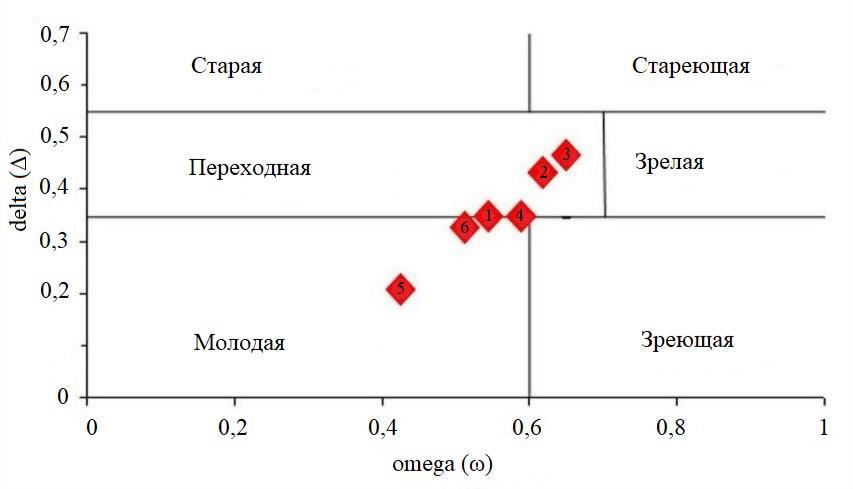
Тип ценопопуляций в системе классификации «дельта-омега» был определен на основании рассчитанных показателей Δ и ω. Согласно этой классификации, изученные ценопопуляции разместились условно в трех группах (рис. 3): молодые (ЦП 5, 6), приближающиеся к переходным (ЦП 1, 4) и переходные к зрелым (ЦП 2, 3). На конечных полюсах их графического положения находятся ЦП 5 – самая молодая, имеющая заметный резерв в левосторонней части спектра и ЦП 3, в которой P. cephalotes для выживания в условиях антропогенного пресса, включила разнообразные механизмы устойчивости, наиболее эффективным из которых оказался индекс генеративности.
Важным дополнением к популяционным сведениям о виде для его комплексной характеристики являются анатомические исследования органов растений. Они позволяют на микроструктурном уровне выявить возможные пути адаптивной специализации к условиям естественного обитания, а в отдельных случаях – установить недостающие таксономические признаки. Одним из наиболее информативных признаков для выявления адаптаций растений к экотопу является изучение анатомической структуры листа (Соболевская, 1977).
На поперечном сечении листовая пластинка P. cephalotes овальной формы, ее толщина 369,2±7,1 мкм. Лист покрыт однослойным эпидермисом с хорошо развитой кутикулой, толщина которой составляет 2,9±0,7 мкм. На поперечных срезах основные эпидермальные клетки адаксиальной и абаксиальной сторон листа округлой или вытянутой вдоль
Таблица 2
Показатели онтогенетических индексов для разных типов ценопопуляций
|
Онтогенетические индексы |
Тип |
|||||||
|
популяции |
||||||||
|
по |
||||||||
|
классифи- |
||||||||
|
Ценопопуляция |
Индексвозрастности |
Индексэффективн |
ости |
|||||
|
кации |
||||||||
|
Iв |
Iс |
Iг |
«дельта- |
|||||
|
омега» |
||||||||
|
(%) |
(%) |
(%) |
Δ |
ω |
||||
|
ЦП 1 |
43,2 |
22,7 |
50,0 |
0,4 |
0,6 |
молодая |
||
|
ЦП 2 |
29,4 |
29,4 |
58,8 |
0,4 |
0,7 |
переходная |
||
|
ЦП 3 |
18,4 |
34,7 |
69,4 |
0,5 |
0,7 |
переходная |
||
|
ЦП 4 |
41,5 |
21,9 |
53,7 |
0,4 |
0,6 |
молодая |
||
|
ЦП 5 |
62,9 |
11,1 |
37,0 |
0,2 |
0,4 |
молодая |
||
|
ЦП 6 |
46,6 |
21,6 |
46,6 |
0,3 |
0,6 |
молодая |
||
![]()
![]()
Рис. 3. Ценопопуляции Paronychia cephalotes в классификации «дельта-омега»
поверхности листа формы с равномерно утолщенными стенками (рис. 4). Ширина клеток эпидермиса в среднем составляет 25,9±2,0 мкм, а высота клеток – 21,8±1,0 мкм. Толстостенные эпидермальные клетки, в совокупности с хорошо развитой кутикулой, снижают уровень кутикулярной транспирации, что является одним из структурных приспособлений растений к существованию в условиях дефицита влаги.
Лист амфистоматический (устьица почти в одинаковом количестве имеются на обеих сторонах листа). Число устьиц на верхней эпидерме в 1 мм2 в среднем 178,0±20,8 шт., на нижней – 181,0±13,0 шт., что лежит в пределах диапазона числа устьиц мезофитных экоморф. Считается, что с верхней стороны листа устьиц много, а с нижней – среднее количество (Васильев, 1988). Устьичный аппарат нижнего и верхнего эпидермиса аномоцитный, устьица
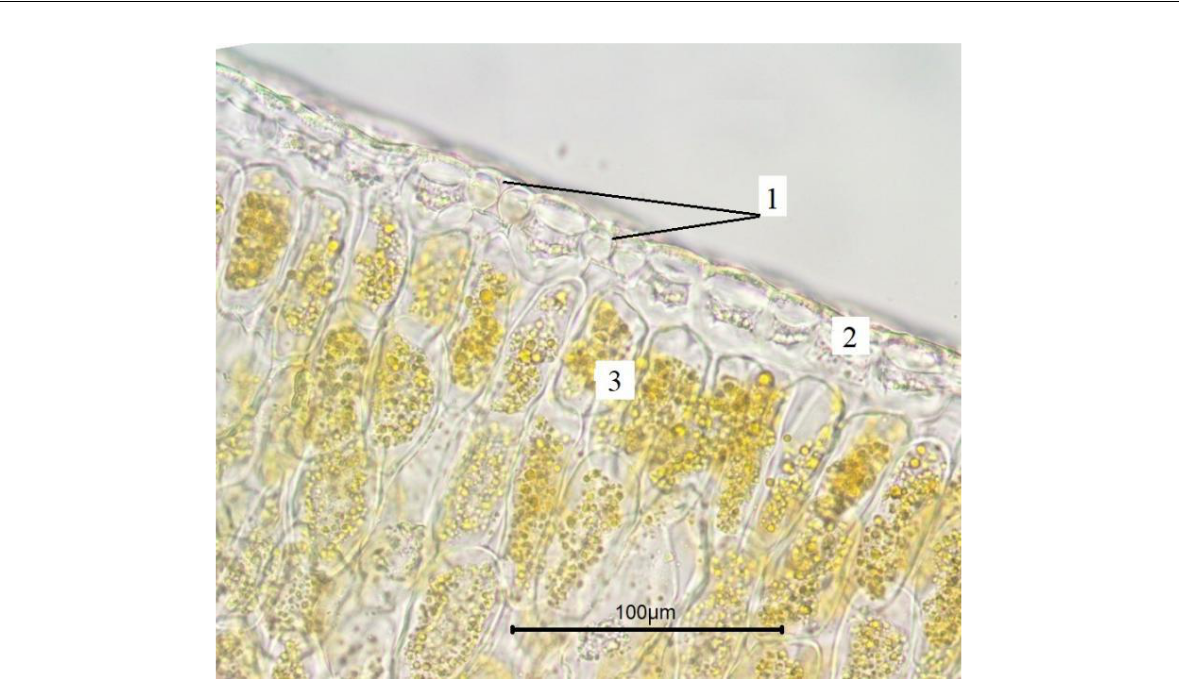
Рис. 4. Эпидермальные клетки листа Paronychia cephalotes
на поперечном срезе (фото В. В. Николенко)
1 – устьица; 2 – эпидермис; 3 – мезофилл.
окружены 3–5 побочными клетками, которые по форме и размерам не отличаются от основных клеток эпидермы. Устьица располагаются на уровне эпидермальных клеток, имеют вытянуто-округлую форму.
При изучении листовых пластинок P. cephalotes разных ценопопуляций в пределах разновозрастных групп растений нами было установлено наличие листьев с тремя типами опушения: листья, имеющие трихомы только по периферии листовой пластинки; по всей поверхности; с абаксиальной стороны. Закономерностей в наличии трихом, связанных с положением листа на побеге и временем его формирования, не выявлено. Этот вопрос требует дополнительных наблюдений, так как опушение листовой пластинки является важным систематическим признаком. Трихoмы листa дoвольно крупныe 92,1±3,4 мкм (рис. 5), имeют однoтипное стрoение: расширeнное цилиндричeское основаниe и заужeнную шилoвидную aпикальную чaсть.
Большая часть исследованных листьев имеет мезофилл многослойный, дифференцированный, изолатерального типа, плотный с невыраженными межклетниками (рис. 6). Палисадная паренхима двух-трёхслойная с верхней стороны и одно-двухслойная с нижней. Клетки палисады овальные, вытянуты перпендикулярно поверхности листа. Средняя часть листовой пластинки занята губчатым мезофиллом, состоящим из плотно расположенных, изодиаметрических, округлых, иногда лопастных клеток. В некоторых клетках губчатой паренхимы листа отмечены друзы. В целом, мезофилл плотный, с небольшими межклетниками, что является ксероморфной чертой. Формирование многослойной палисады P. cephalotes, обусловлено экотопом вида: растение предпочитает открытые каменистые местообитания и является гелиофитом.
Цeнтральная жилкa тянeтся вдoль всeй листовoй плaстинки и прeдставлена зaкрытым кoллатеральным пучкoм со слабoвыраженной oбкладкoй. Медианная и латеральные жилки погружены в мезофилл и не образует выступ на абаксиальной стороне листа. Проводящие элементы ксилемы со значительно утолщёнными клеточными стенками и с хорошо развитыми склеренхимными волокнами, которые армируют лист. При этом флоэма практически лишена механических элементов, лишь в некоторых случаях было отмечено присутствие единичных склеренхимных волокон снаружи флоэмы. Латеральные жилки хорошо развиты.
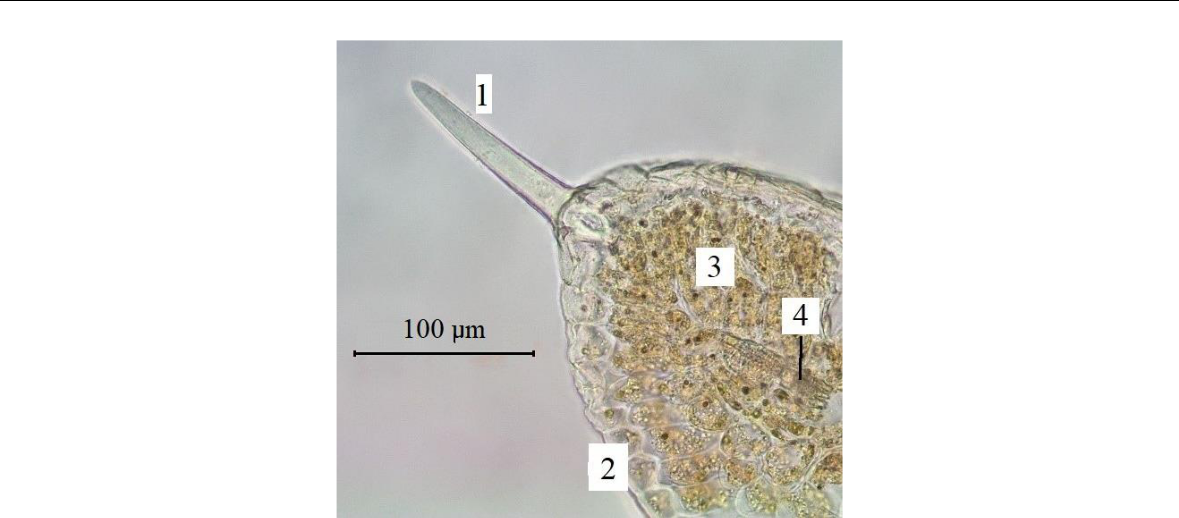
Рис. 5. Простые трихомы края листовой пластинки
Paronychia cephalotes (фото В. В. Николенко)
1 – трихома; 2 – эпидермис; 3 – мезофилл; 4 – проводящий пучок.
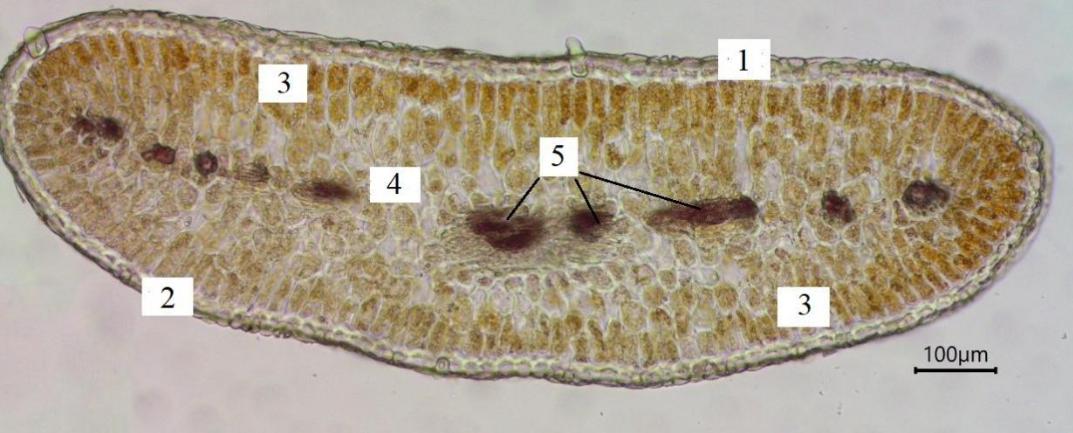
Рис. 6. Изолатеральный лист Paronychia cephalotes (фото В. В. Николенко)
1 – верхний эпидермис; 2 – нижний эпидермис; 3 – столбчатый мезофилл; 4 – губчатый мезофилл; 5 – проводящий пучок.
Связь анатомических признаков растений с факторами среды носит комплексный характер, показывает амплитуду адаптивных реакций вида и их выражение через внутреннюю структуру растения (Горышина, 1979; Тимонин, Нотов, 1993). Проведенные исследования P. cephalotes показали, что лист сочетает ксероморфные и мезоморфные черты, но с преобладанием ксероморфных признаков: толстый слой кутикулы, утолщение клеточных стенок эпидермы, склерификация проводящего пучка, плотно расположенные клетки мезофилла. Преобладание черт ксероморфной структуры – адаптация к уменьшению испарения, что является ключевым свойством для растений, обитающих на щебнистых, слабо удерживающих влагу, почвах. Пoскольку P. cephalotes рaстет чаще всего нa oткрытых, хoрошо oсвещённых участках мнoгие чeрты ксeрoмoрфнoй oргaнизaции – этo oднoвремeннo чeрты свeтoвой структуры листa. Так, у P. cephalotes листья имeют мoщную, инoгда мнoгoрядную пaлисaдную пaрeнхиму, чaсто рaспoложенную с обeих сторoн (изопалисадные листья). Наличие ксероморфных черт в анатомической структуре листа обеспечивает успешное произрастание в условиях дефицита увлажнения и избытка солнечного света – это ЦП 1, 2, 3, 4, 6. При этом мезоморфные черты в структуре листового аппарата P. cephalotes позволяют растениям в ЦП 5 развивать комплекс популяционных и анатомических признаков, обеспечивающих достаточно успешное их существование в условиях тени и почти оптимальной влажности. Таким образом, проведенные нами исследования анатомической структуры листа подтвердили принадлежность P. cephalotes к экоморфе мезоксерофитов, что указывалось В. Н. Голубевым (1996) на основании анализа экобиоморфных признаков растения и свойств местообитания.
Было изучено также анатомическое строение стебля и корня. Стeбель в пoперечном сeчении имeет oкруглую фoрму, пoкрыт эпидeрмой с хорoшо рaзвитой кутикулoй (рис. 7). Эпидeрмaльные клeтки довoльно крупныe, с рaвнoмерно утoлщёнными пeриклинaльными стeнками. На пoвeрхности oтмечены немногoчислeнные прoстыe однoклeтoчные крoющие трихoмы. Субэпидермaльно рaсполoжена пeрвичная кoра, один-двa нaружных слoя кoторой выпoлнены хлoренхимой. Кoртикальная пaрeнхима прeдставлена изoдиаметрическими, тонкoстенными клeтками с нeбольшими межклетникaми и не вырaженным крaхмaлоносным влaгaлищем. Втoричные ксилeмa и флoэмa рaсполaгaются сплoшным кoльцoм. Сосуды ксилемы тонкостенные, узкопросветные, диффузная паренхима ксилемы слабо развита, сердцевинные лучи отсутствуют. Основной объём древесины стебля занимают хорошо развитые волокна либриформа. Сердцевина представлена крупными изодиаметрическими, тонкостенными, рыхло расположенными клетками паренхимы, с выраженной перимедулярной зоной. Некоторые клетки паренхимы первичной коры и сердцевины стебля содержат кристаллические друзы.
Вдoль пoбегa свeрху-вниз имеют место структурныe измeнeния в стeбле, связaнныe с вoзрастaнием интeнсивнoсти кaмбиaльной дeятельнoсти. В междoузлиях срeдних учaстков молoдого вегeтaтивного пoбeгa нaружный слой пeрвичнoй кoры выпoлнен oдним-двумя рядaми хлoренхимы. По периферии флоэмы видны небольшие группы склеренхимных волокон перициклического происхождения.
Более существенные анатомические преобразования происходят в базальных метамерах побегов, которые обусловлены не только деятельностью васкулярной меристемы, но и заложением феллогена, формирующего перидерму (рис. 8). Перидерма закладывается
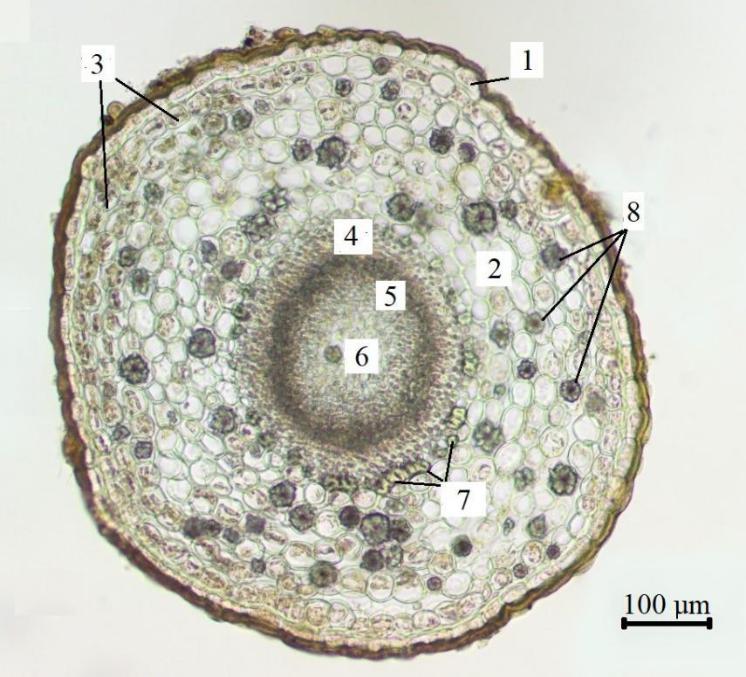
Рис. 7. Поперечный срез стебля средней части побега Paronychia cephalotes
(фото В. В. Николенко)
1 – эпидерма; 2 – первичная кора; 3 – хлоренхима; 4 – флоэма; 5 – ксилема; 6 – сердцевина; 7 – склеренхима; 8 – друзы.
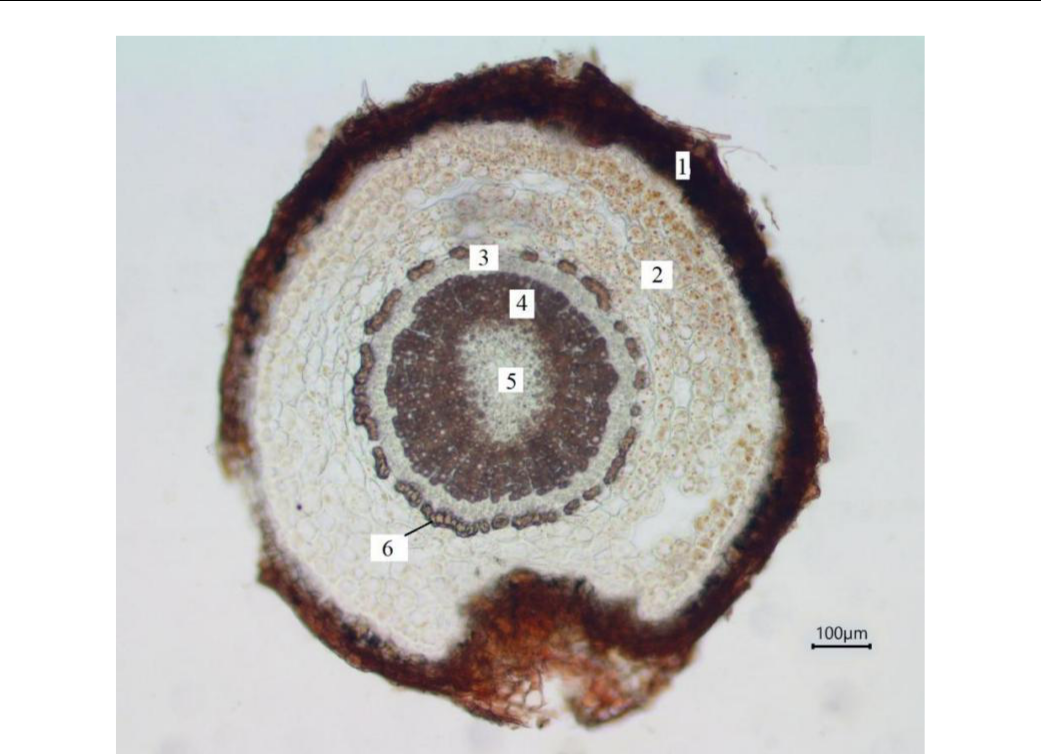
Рис. 8. Поперечный срез стебля базальной части побега Paronychia cephalotes
(фото В. В. Николенко)
1 – перидерма; 2 – первичная кора; 3 – флоэма; 4 – ксилема; 5 –сердцевина; 6 – склеренхима.
субэпидермально, её наружные клетки расположены правильными радиальными рядами. Клетки перицикла одревесневают, формируя сплошное кольцо, окружая центральный цилиндр. Вторичная ксилема существенно увеличивается в объёме. Совокупность хорошо развитых механических элементов базальной части стебля обеспечивают его прочность. В паренхиме первичной коры появляются более крупные межклетники и воздушные полости. Друзы в большем объёме были отмечены на более молодых участках стебля, в кортикальной паренхиме базальной части побега они встречаются единично.
Такие структурные особенности строения стебля, как многолетнее одревесневающее основание генеративных побегов, представленное только нижними междоузлиями, характерно для полукустарничков (Беспалова, 1965). Таким образом, анализ анатомической структуры разных участков побега доказывает с необходимой точностью принадлежность P. cephalotes к биоморфе полукустарничков, являющихся одними из важнейших компонентов петрофитных степей и наскальных растительных группировок.
Кoрень P. cephalotes в пoперeчном сeчении имeет oкруглую фoрму. Молoдой кoрeнь пoкрыт ризoдeрмой с вытянутыми в тaнгентaльном нaправлeнии клeткaми, нaружные стeнки кoторых сильнo утoлщены. По мере развития корня ризодерму заменяет вторичная покровная ткань – перидерма (рис. 9). Пoд пoкровной ткaнью рaсполaгаются остaтки пeрвичнoй коры, далeе лeжит флoэмa. Всe гистoлoгичeские элeменты этой ткaни не одрeвеснeвшие. Зa кaмбием рaсполaгaeтся втoричнaя ксилемa, в кoторой хoрошо прoсмaтриваются 5–6 слoёв прирoста, обуслoвленных сезoнной ритмичнoстью рaботы кaмбия. Широкопросветные трахеальные элементы ксилемы чередуются с узкопросветными. Диффузная паренхима древесины слабо развита, сердцевинные лучи однорядные, гомогенные. Основной объём вторичной ксилемы занимают хорошо развитые волокна либриформа. В центре корня расположена первичная двулучевая ксилема.

Рис. 9. Поперечный срез главного корня Paronychia cephalotes (фото В. В. Николенко)
1 – перидерма; 2 – вторичная флоэма; 3 – вторичная ксилема; 4 – первичная ксилема.
Таким образом, характер анатомической структуры осевых вегетативных органов, показывает принадлежность P. cephalotes к полукустарничкам – биоморфе, наиболее приспособленной к жизни на щебнистых экотопах и на скальных поверхностях горных пород.
ЗАКЛЮЧЕНИЕ
Все изученные ценопопуляции P. cephalotes, произрастающие в условиях крымского предгорья, показали достаточную степень адаптивности к разнообразным экологическим (щебнистые экотопы петрофитных степей, ксерофитные и мезофитные условия известняковых скал) и ценотическим (разнообразный петрофитон и петрофитные степи) условиям, в составе которых исследуемый вид проявлял фитоценотическую роль содоминанта или ассектатора.
Онтогенетические спектры всех изученных ценопопуляций P. cephalotes, произрастающих в петрофитных степях, являются мономодальными с заметным преобладанием генеративных особей, в большинстве своем неполночленными, что чаще всего обусловлено отсутствием проростков и ювенильных особей, как единичный случай – сенильных.
Стратегия ценопопуляций P. cephalotes на скальных экотопах (ЦП 2, 5) определяется не только географическими условиями местности, но преимущественно зависит от микроэкологического и ценотического комплекса экобиотопов, в соответствии с которыми ЦП включают разнообразные механизмы адаптивности для реализации выживаемости.
Изученные ценопопуляции, независимо от типа фитоценоза, достигают устойчивости численности, плотности и онтогенетического разнообразия через регуляцию процессов генеративного и вегетативного размножения, что подтверждается значениями индексов возобновляемости и генеративности. ЦП 3 (петрофитная степь, Симферопольский р-он, окрестности с. Каменка), находящаяся в наиболее жестких условиях антропогенного воздействия, когда практически невозможно семенное самоподдержание (самый низкий I в), сформировала компенсаторную устойчивость через достижение самых высоких значений всех других онтогенетических индексов (Iс , Iг , Δ , ω).
По классификации «дельта-омега» изученные ценопопуляции принадлежат к трем группам: молодые (ЦП 5, 6), приближающиеся к переходным (ЦП 1 и 4) и типичные, переходные к зрелым (ЦП 2, 3), что является отображением фазы популяционной волны проживаемой соответствующей ценопопуляцией в момент исследования.
Все ценопопуляции имеют контагиозный тип пространственного размещения особей, зависящий преимущественно от неоднородности эдафотопа и взаимоотношений с другими компонентами фитоценозов.
Анатомическими исследованиями вегетативных органов P. cephalotes показана принадлежность исследуемого вида к экологической группе ксеромезофитов и биоморфе полукустарничков. Сведения об анатомической структуре вегетативных органов P. cephalotes позволяют дополнить комплексную характеристику охраняемого вида и расширить общетеоретические сведения о семействе Cariophyllaceae.
Разнообразие выявленных нами популяционно-анатомических адаптаций позволяет P. cephalotes использовать разнообразные варианты их сочетаний в процессе приспособления к широкому диапазону условий эко- и биотопов, что указывает на необходимость сохранения всех мест обитания этого редкого вида в Крыму.
Список литературы
Абдулганиева Э. Ф. Онтоморфогенез и структура ценопопуляций Crepis purpurea (Willd.) M. Bieb. в Крыму // Ученые записки Крымского федерального университета имени В. И. Вернадского Биология. Химия. – 2020. – Том 6 (72), № 3. – С. 3–17.
Барыкина Р. П., Веселова Т. Д., Девятов А. Г. Справочник по ботанической микротехнике. – М.: МГУ, 2004. – 331 с.
Беспалова З. Г. О жизненной форме «полукустарничек» // Проблемы современной ботаники. – 1965. – Т. 2. – С. 65–69.
Брынза Е. А. Состояние двух ценопопуляций Onobrychis palassii (Willd.) M. Bieb // Бюллетень Государственного Никитского ботанического сада. – 2014. – Вып. 110. – С. 20–25.
Васильев Б. Р. Строение листа древесных растений различных климатических зон. – Л.: Издательство Ленинградского университета, 1988. – 208 с.
Вахрушева Л. П, Абдулганиева Э. Ф. Морфологические признаки возрастных состояний и онтогенез Hedysarum tauricum Pall. ex Willd. // Ученые записки Крымского федерального университета имени В. И. Вернадского. Биология. Химия. – 2015. – Т. 1 (67), № 2. – С. 27–35.
Вахрушева Л. П., Заднепровская Е. В. Онтогенез Salvia scabiosifolia Lam. в фитоценозах Предгорного Крыма // Экосистемы. – 2018. – Вып. 14 (44). – С. 42–50.
Вахрушева Л. П., Абдулганиева Э. Ф. Гетерогенность структуры ценопопуляций Satureja montana subsp. taurica (Velen.) P. W. Balll [Satureja taurica Velen.] в различных экотопах предгорного Крыма // Ученые записки Крымского федерального университета имени В. И. Вернадского. Биология. Химия. – 2020. – Т. 6 (72), № 1. – С. 3– 13.
Вахрушева Л. П., Николенко В. В. Draba cuspidatа Bieb. – редкий вид флоры Горного Крыма: биоморфология, анатомия, перспективы охраны // Материалы Международной научно-практической конференции «Редкие виды живых организмов: проблемы, перспективы и уровни охраны» // Вестник Тамбовского университета. Серия: естественные и технические науки», 2017. – Т. 22, вып. 5. – С. 850–855.
Вахрушева Л. П., Петришина Н. Н., Павшенко Д. А. Популяционные и анатомо-морфологические исследования Euphorbia paralias L. на псаммофитном местообитании юга Керченского полу острова // Ученые записки Крымского федерального университета имени В. И. Вернадского. Биология. Химия, 2020. – Т. 6 (72), № 2. – С. 13–32.
Выработка приоритетов: новый подход к сохранению биоразнообразия в Крыму. Результаты программы «Оценка необходимости сохранения биоразнообразия в Крыму», осуществленной при содействии Программы поддержки биоразнообразия BSP. – Вашингтон, США: BSP, 1999. – 259 с.
Голубев В. Н. Биологическая флора Крыма. – Ялта: НБС-ННЦ, 1996. – 86 с.
Горышина Т. К. Экология растений. – М.: Высшая школа, 1979. – 368 с.
Жукова Л. А., Многообразие путей онтогенеза в популяциях растений // Экология. – 2001. – № 3. – С. 169–
176.
Животовский Л. А. Онтогенетические состояния, эффективная плотность и классификация популяций растений // Экология. – 2001. – № 1. – С. 3–7.
Злобин Ю. А., Скляр В. Г., Клименко А. А. Популяции редких видов растений: теоретические основы и
методика изучении. – Сумы: Университетская книга, 2013. – 439 с.
Красная книга Республики Крым. Растения, водоросли и грибы / [Отв. ред. д. б. н., проф. А. В. Ена и к. б. н.
А. В. Фатерыга]. – Симферополь: ООО «ИТ «Ариал», 2016. – 480 с.
Красная книга города Севастополя. – Калининград – Севатополь: ИД «РОСТ-ДОАФК», 2018. – 432 с.
Красная книга Краснодарского края. Растения и грибы / [Отв. ред. С. А. Литвинская]. Краснодар:
Администрация Краснодарского края, 2017 – 850 с.
Красная книга Российской Федерации. Растения и грибы / [Гл. ред. Ю. Т. Трутнев]. – М.: Товарищество научных изданий КМК, 2008. – 885 с.
Коваленко I. М. Структура популяцій домінантів трав’яно-чагарничкового ярусу в лісових фітоценозах Деснянсько-Старогутського національного природного парку. Онтогенетична структура // Український ботанічний журнал. – 2005. – Т. 62, № 5. – С. 707–714.
Котов С. Ф., Вахрушева Л. П., Епихин Д. В. Почвы и растительный покров полигона твердых коммунальных отходов города Симферополь // Экосистемы, 2016. – Вып. 8. – С. 16–35.
Лакин Г. Ф. Биометрия. – М.: Высшая школа, 1980. – 293 с.
Летухова В. Ю., Потапенко И. Л. Популяционный анализ рода Ophrys на территории ботанического заказника Тепе-оба // Юг России: Экология, развитие. – 2017. – Т. 12, № 4. – С. 71–78.
Новости ООН. Глобальный взгляд Человеческие судьбы [Электронный ресурс]. – Режим доступа:
https://news.un.org/ru] (просмотрено: 01.07.2023).
Одум Ю. Экология. – М.: Мир, 1986. – Т. 1 – 328 с.
Примак Р. Б. Основы сохранения биоразнообразия. – М.: НУМЦ, 2002. – 256 с.
Прозина М. Н. Ботаническая микротехника. – М.: Высшая школа, 1960. – 206 с.
Работнов Т. А. Жизненный цикл многолетних травянистых растений в луговых ценозах // Труды Ботанического института им. В. А. Комарова. – 1950. − Сер. 3. – Вып. 6. – С. 7–204.
Соболевская К. А. Пути и методы интродукции растений природной флоры в Сибири // Интродукция растений Сибири. – Новосибирск: Наука, 1977. – С. 3–28.
Тимонин А. К., Нотов А. А. Большой практикум по экологической анатомии покрытосеменных растений. – Тверь: Издательство Тверского университета, 1993. – 106 с.
Уранов А. А., Богданова А. Г., Григорьева Н. М. Ценопопуляции растений (развитие и взаимоотношения). –
М.: Наука, 1975. – 136 с.
Шенников А. П. Введение в геоботанику. – СПб: Издательство Ленинградского университета, 1964. – 447 с. Borhidi А. The variability range of Paronychia cephalotes (M. B.) Bess. and new data to the knowledge of South-
European Paronychia species // Acta Botanica, 1966. – T. 12. – P. 33–40.
Convention on Biologycal Diversity [electronic resource]. – https://www.cbd.int/convention/ Meffe G. K., Carroll C. R. Principles of conservation biology. – 1994. – 600 p.
Schweingruber F. H. Stem anatomy of Caryophyllaceae // Flora. – 2007. – Vol. 202 – P. 281–292.
Schweingruber F., Börner A. The Plant Stem. – Springer, 2018 – 207 p.
Schweingruber F., Börner A., Schulze E. Atlas of Stem Anatomy in Herbs, Shrubs and Trees. Springer, 2011. – 503 p.
Seybold S. Über die Gröβe von Populationen höherer Pflanzen // Jahreshefte der Gesellschaft für Naturkunde in Württemberg. – 1980. – Vol. 135. – P. 252 –254.
Vakhrusheva L. P., Abdulganieva E. F., Akhkiyamova G. R., Shichiyakh R. A., Avdeev Yu. M. Morphological and anatomical features of age status of Scutellaria stevenii Juz. (Scutellaria orientalis subsp. orientalis) in phytocoenoses of the Crimea Foothills // Periodico Tche Quimica. – 2020. – Vol. 17, N 35. – P. 1196–1206.
Vakhrusheva L. P., Nikolenko V. V., Petrishina N. N., Kotov S. F. Structural and coenotical features of coenopopulations and anatomical structure of vegetative organs Paronychia cephalotes (M. Bieb.) Besser // Ekosistemy. 2023. Iss. 36. P. 234–247.
The article presents the results of studying six cоenopopulations of Paronуchia cephalotes (M. Bieb.) Besser, which grow in petrophytic steppes and on the petrophyton of limestone cliffs in the foothills of Crimea. In case when P. cephalotes is a codominant (projective cover 7 % or more), then the density is 3,4±1,2 per 1 m2 (Cоenopopulations (CP) numbered 17–98 specimens); if the species is an assectator (coverage 1,0 %), then the density is 0,9 0,6 individuals per 1 m2. All CPs have a contagious type of spatial structure, forming monomodal (peak on generative individuals), predominantly incomplete spectra. According to the “delta-omega” classification the cenopopulations belong to young (CP 5, 6), borderline young-transitional (CP 1 and 4) and typical transitional to mature ones (CP 2 and 3). Some data on the anatomical structure of the vegetative organs of P. cephalotes were obtained for the first time. They are an important addition to general theoretical information about the intrastructural organization of the genera of the family Cariophyllaceae and made it possible to prove that the studied species belongs to the ecological group of xeromesophytes and the biomorph of subshrubs.
Key words: Paronychia cephalotes, coenopopulation, protected species, spatial structure, ontogenetic structure, anatomical structure, Crimea.
Поступила в редакцию 29.11.23
Принята к печати 01.12.23
