
| Экосистемы, 37: 89–95 (2024) | https://ekosystems.cfuv.ru |
| УДК 574.45+582.28 | DOI: 10.29039/2413-1733-2024-37-89-95 |
Процессы микогенного ксилолиза в искусственных древесных насаждениях степной зоны Оренбургской области (Южное Предуралье)
Оренбургский государственный университет
Оренбург, Россия
safonovmaxim@yandex.ru
Грибы-макромицеты – важнейший компонент системы редуцентов лесных экосистем. Процесс микогенной деструкции древесины зависит от многих факторов, а это в свою очередь, влияет на функционирование лесных экосистем. Исследование процессов микогенного ксилолиза в Южном Предуралье (Оренбургская область) было начато в 2000 году, однако эксперименты проводились исключительно в пределах лесостепной части области и затрагивали древостои естественного происхождения. При этом большая часть региона расположена в пределах степной зоны, что определяет актуальность аналогичных исследований в центральных и южных районах области
- древостоях искусственного происхождения. Эксперимент проводился в 2019–2022 году; образцы древесины березы, сосны, вяза и клена были заложены на срок 3 года Проведенные исследования показали, что в пределах степной зоны Южного Предуралья скорость микогенной деструкции древесины варьирует. Максимальная скорость разрушения характерна для веточного отпада вяза, березы и сосны, наименьшая – для отпада клена. Более широкое варьирование показателей скорости разложения было характерно для образцов древесины сосны и березы. Скорость разложения образцов сосны существенно варьирует в зависимости от локации, в то время как для древесины березы показатели более однообразны. Сопоставление полученных материалов с полученными ранее данными о скорости микогенного ксилолиза в лесах лесостепной части региона показывает, что скорость разложения древесины в искусственных насаждениях несколько ниже аналогичного показателя в естественных лесах; это особенно заметно в отношении деструкции древесины сосны. Таким образом, интенсивность микогенного ксилолиза в искусственных насаждениях степной зоны Южного Предуралья относительно высока и вполне сопоставима с таковой в естественных насаждениях. Это свидетельствует о эффективной деятельности системы редуцентов в искусственных насаждениях.
Ключевые слова: грибы-макромицеты, микогенный ксилолиз, искусственные насаждения, Южное Предуралье, Оренбургская область.
ВВЕДЕНИЕ
Устойчивое существование биогеоценозов обеспечивается сбалансированными процессами продукции и деструкции. Продукционные характеристики биогеоценоза определяется многими факторами, такими как количество поступающей солнечной радиации, структурно-функциональные характеристики фитоценоза, деятельностью травоядных животных и так далее. Деструкционные процессы, обеспечивающие возврат в круговорот биогеоценоза вещества и энергии, реализует система редуцентов, в состав которой входят бактерии и грибы. В определенной мере к деструкторам, точнее – к измельчителям, относятся беспозвоночные, однако именно грибы и бактерии способны переработать органику в более легко усваиваемую форму.
- лесных экосистемах важнейшим компонентом системы редуцентов являются грибы, в особенности макромицеты. Среди них есть гумусовые и подстилочные сапротрофы, копротрофы и ксилотрофы. Большое влияние на состояние древостоев оказывают ксилотрофы – дереворазрушающие грибы, которые обеспечивают разложение целлюлозы и лигнина пней и отмерших ветвей за счет специфической гарнитуры ферментов. Они также регулируют структуру древостоев за счет видов, поселяющихся на ослабленных деревьях, формирующих гнили и приводящих к выпадению этих деревьев из насаждений.
Роль дереворазрушающих грибов в круговороте вещества в лесных экосистемах очень важна, так как большая часть углерода депонируется именно в древесине и от деятельности ксилотрофных грибов зависит – насколько быстро этот углерод высвободится.
Микогенная деструкция древесины – процесс, зависящий от многих факторов. От скорости и эффективности этого процесса зависят многие аспекты функционирования лесных экосистем. Исследование этого процесса в России происходит достаточно давно; в настоящее время большое внимание к этой теме связано с проблемами контроля углерода, как одного из важнейших факторов глобальной трансформации климата (Волчатова, 2007; Кузнецов, 2009; Шорохова, 2009; Казарцев, 2014; Мухин, 2015; Стороженко, 2016; Диярова, 2020; Mukhin, 2007). Все исследования позволяют с определенной долей уверенности говорить о варьировании характеристик этого процесса в зависимости от родовой принадлежности субстрата, особенностей организмов-деструкторов, длительности процесса разложения, а также в зависимости от типа леса и его положения в комплексном экологическом градиенте.
Видовой состав деструкторов является очень важным фактором, определяющим ход процесса разложения из-за различий в составе ферментного комплекса видов, а также интенсивности выработки этих ферментов. Влияние этого аспекта физиологии дереворазрушающих грибов на скорость разложения древесины было неоднократно доказано экспериментально (Степанова, Мухин, 1979; Progar, 2000). Ход деструкции также зависит от размеров субстрата. Чем больше размер субстрата, тем больше требуется времени на его разложение (Boddy, 1995). Также на деструкционные процессы оказывают влияние особенности среды, то есть биотопа – определенный тип леса, положение в пределах определенной природно-климатической зоны, уровень антропогенной нагрузки и так далее (Progar, 2000; Mukhin, 2007 и др.).
Исследование процессов микогенного ксилолиза в Южном Предуралье (Оренбургская область) было начато нами в 2000 году (Сафонов, 2006; Сафонов, Булгаков, 2013) и позволило получить вполне репрезентативные данные о характеристиках процесса, однако эксперименты проводились исключительно в пределах лесостепной части области и затрагивали древостои естественного происхождения. Так как большая часть региона расположена в пределах степной зоны, мы сочли интересным провести аналогичные исследования в центральных и южных районах области. Естественные леса в степной зоне Южного Предуралья не многочисленны и представлены преимущественно пойменными лесами (вязовники, тополевники, кленовники, ивняки), березовыми и осиновыми лесами, произрастающими по балкам, широколиственными лесами на склонах возвышенности Общий Сырт. Лесистость большинства районов составляет менее 4 %.
-
- степных районах значительная часть лесных насаждений представлена защитными лесополосами. Лесополосы и другие искусственные древесные насаждения являются качественно своеобразным типом антропогенных экосистем, которые уже несколько веков являются неотъемлемой частью общего облика природы, в особенности в условиях аридного
- субаридного климата (Сафонов и др., 2013). Распространение искусственных насаждений определяется тем, что степные и полупустынные регионы с низкой естественной лесистостью нуждаются в значительном количестве лесных насаждений разного типа для повышения эффективности сельскохозяйственного производства, повышения комфортности условий обитания человека, снижения антропогенных (техногенных) нагрузок на природно-территориальные комплексы.
Исходя из природной специфики региона, мы поставили перед собой цель – изучить скорость микогенного ксилолиза в искусственных лесных насаждениях степной части Южного Предуралья и сопоставить полученные результаты с аналогичными данными из лесостепных районов.
МАТЕРИАЛ И МЕТОДЫ
Оценка скорости биологического разложения древесины проводилась по методике, основанной на закладке в подстилку образцов ветвей диаметром 1–5 см, длиной 15–20 см известной массы. По убыли массы образцов за определенный период времени оценивали величину биологического разложения (Степанова, Мухин, 1979).
Эксперимент проводился в 2019–2022 годах. Образцы древесины были заложены на срок 3 года в следующих локациях: посадка сосны в Акбулакском районе (окрестности села Григорьевка), Платовская лесная дача (Новосергиевский район), Шубарагашская лесная дача (Соль-Илецкий район), окрестности села Шумаево (Ташлинский район), березовая посадка у села Красноперовск (Саракташский район) (рис. 1).
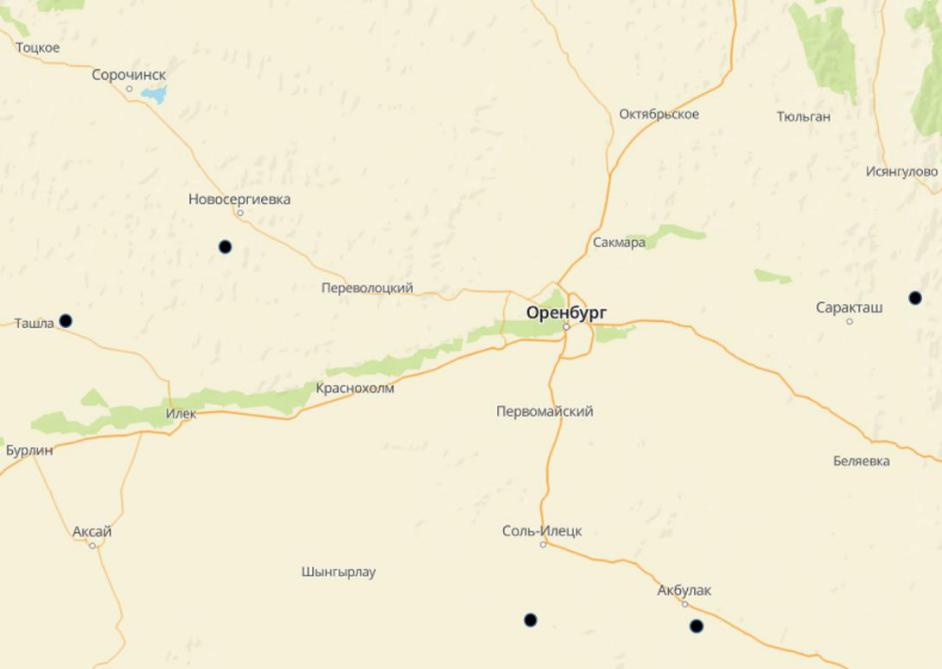
Рис. 1. Локации проведения эксперимента
Ниже приводятся описания насаждений, в которых производилась закладка образцов. Категорию состояния деревьев оценивали глазомерно. Для оценки относительного жизненного состояния (ОЖС) насаждений была принята методика В. А. Алексеева (1990).
Сосновые насаждения: Новосергиевский район, село Рыбкино, «Платовская лесная дача» (ОЖС 75,1 %, высота: 24 м, возраст 60 лет, полнота 0,2, в подлеске клен остролистный, клен американский, вяз шершавый, клен татарский); Соль-Илецкий район, Шубарагашская лесная дача (ОЖС 41,6 %, высота: 10 м, возраст 36 лет, полнота 0,4, в подлеске смородина золотистая, крушина ломкая); Акбулакский район, окрестности села Григорьевка (ОЖС 36,1 %, высота: 10 м, возраст 46 лет, полнота 0,5, подлесок отсутствует).
Березовые насаждения: Саракташский район, село Красноперовск (ОЖС 72,3 %, высота 12 м, возраст 25 лет, сомкнутость крон 0,85, полнота 0,5, в подлеске черемуха, клен остролистный, рябина, шиповник, малина); Новосергиевский район, село Рыбкино, «Платовская лесная дача» (ОЖС 85,3 %, высота 20 м, возраст 40 лет, сомкнутость крон 0,8, полнота 0,5, в подлеске: робиния ложноакация, шиповник коричный, клен остролистный, черемуха, вяз гладкий); Соль-Илецкий район, Шубарагашская лесная дача (ОЖС 69,3 %, высота 10–12 м, возраст 18 лет, сомкнутость крон 0,65, полнота 0,3, подлесок отсутствует).
Посадка клена остролистного: Новосергиевский район, село Рыбкино, «Платовская лесная дача» (ОЖС 61,7 %, высота 20 м возраст 20 лет, сомкнутость крон 0,7, полнота 0,5, в подлеске жимолость татарская).
Посадка вяза гладкого: Ташлинский район, окрестности села Шумаево (ОЖС 71,3 %, высота 20 м, возраст 25 лет, сомкнутость крон 80 %, полнота 0,3, в подлеске смородина золотистая и жимолость татарская).
Было заложено и проанализировано по 50 образцов древесины каждого вида. В общей сложности было заложено и проанализировано 400 образцов. Потеря массы рассчитывалась как разница между исходным весом условно сухого образца в и итоговым весом условно сухого образца (высушивание проводилось в сухожаровом шкафу в течение 4 часов при температуре 104 °С). При анализе данных по каждой выборке определялись минимальные, максимальные показатели, среднее арифметическое и стандартная ошибка, дисперсия выборки.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Проведенные исследования показали, что в пределах степной зоны Южного Предуралья характеристики процесса микогенной деструкции древесины варьируют.
На опытных образцах древесины в ходе эксперимента отмечалось формирование плодовых тел ограниченного числа видов, таких как Cylindrobasidium laeve (Pers.) Chamuris,, Datronia stereoides (Fr.) Ryvarden, Gloeoporus taxicola (Pers.) Gilb. & Ryvarden, Irpex lacteus (Fr.) Fr., Postia hibernica (Berk. & Broome) Julich, Schizophyllum commune Fr. Базидиомы образовались лишь на 6,75 % образцов.
Большая часть образцов имела признаки белой гнили; 26 % образцов сосны имели признаки деструктивной гнили; 30,25 % образцов имели признаки повреждения насекомыми (особенно образцы древесины клена и вяза).
Результаты опыта показали, что в искусственных насаждениях Южного Предуралья наблюдается изменение скорости деструкции древесины в зависимости от ее принадлежности к тому или иному роду древесных растений.
Максимальная скорость разрушения характерна для веточного отпада вяза, березы и сосны, наименьшая – для отпада клена (табл. 1). Более широкое варьирование показателей скорости разложения было характерно для образцов древесины сосны и березы.
Таблица 1
Скорость разложения веточного отпада в лесах Южного Предуралья
(потеря массы за трехлетний период) (n=50)
| Древесина | Локация | Потеря массы, % | ||
| M±m | Lim | σ2 | ||
| Платовская лесная дача | 25,5±9,3 | 6,6–39,6 | 87,1 | |
| Сосна | ||||
| Шубарагашская лесная дача | 21,3 ±10,9 | 5,2–39,9 | 118,1 | |
| (Pinus silvestris) | ||||
| Акбулакский район | 16,63±6,3 | 7,2–28,8 | 39,7 | |
| (с. Григорьевка) | ||||
| Саракташский район | 26,8±8,5 | 8,2–42,0 | 72,5 | |
| Береза | (с. Красноперовск) | |||
| Платовская лесная дача | 24,7±9,8 | 9,1–39,9 | 95,8 | |
| (Betula pendula) | ||||
| Шубарагашская лесная дача | 22,0±9,6 | 6,2–38,9 | 90,5 | |
| Клен | ||||
| остролистный | Платовская лесная дача | 17,7 ±5,9 | 7,4–29,6 | 35,1 |
| (Acer platanoides) | ||||
| Вяз шершавый | Ташлинский район | 23,1±8,1 | 9,1– 41,4 | 65,5 |
| (Ulmus laevis) | (с. Шумаево) | |||
Графический анализ распределения потерь массы образцов (рис. 2) показал, что скорость разложения образцов сосны существенно варьирует в зависимости от локации, в то время как для древесины березы показатели более однообразны.
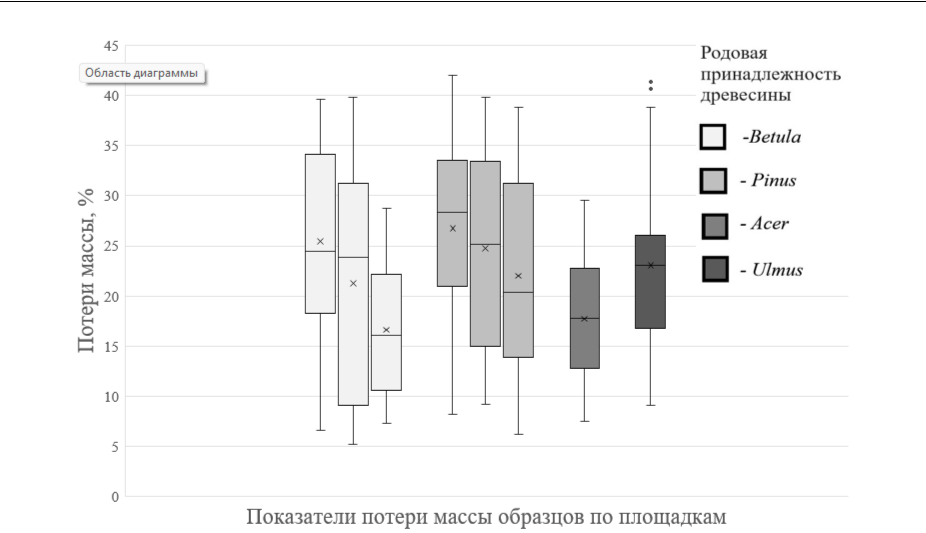
Рис. 2. Варьирование показателей потерь массы образцов по родовой принадлежности древесины
Крестик – среднее арифметическое значение по выборке; черта посередине – медиана по выборке; отдельные точки – значения, сильно отличающиеся от основного массива выборки.
Сопоставление полученных материалов с полученными ранее данными о скорости микогенного ксилолиза в лесах лесостепной части региона (Сафонов, 2006) показывает, что скорость разложения древесины в искусственных насаждениях несколько ниже аналогичного показателя в естественных лесах; это особенно заметно в отношении деструкции древесины сосны (табл. 2). При этом в лесостепной части региона несколько выше дисперсия этого показателя.
Таблица 2
Скорость разложения веточного отпада в лесах Южного Предуралья (потеря массы за трехлетний период) (n=50)
| Древесина | Потеря массы, % | |||||
| M±m | Lim | σ2 | ||||
| Лесостепная зона | ||||||
| Сосна (Pinus silvestris) | 31,6±2,6 | 10,3–55,3 | 131,7 | |||
| Береза (Betula pendula) | 25,4±2,4 | 9,1–45,0 | 102,0 | |||
| Искусственные насаждения степной зоны | ||||||
| Сосна (Pinus silvestris) | 21,1±9,7 | 5,2–39,9 | 93,7 | |||
| Береза (Betula pendula) | 24,5±9,4 | 6,2–42,0 | 89,0 | |||
Сравнение полученных нами данных с аналогичными материалами из других регионов показывает продолжение тренда к снижению скорости микогенного ксилолиза в широтном градиенте (рис. 3).
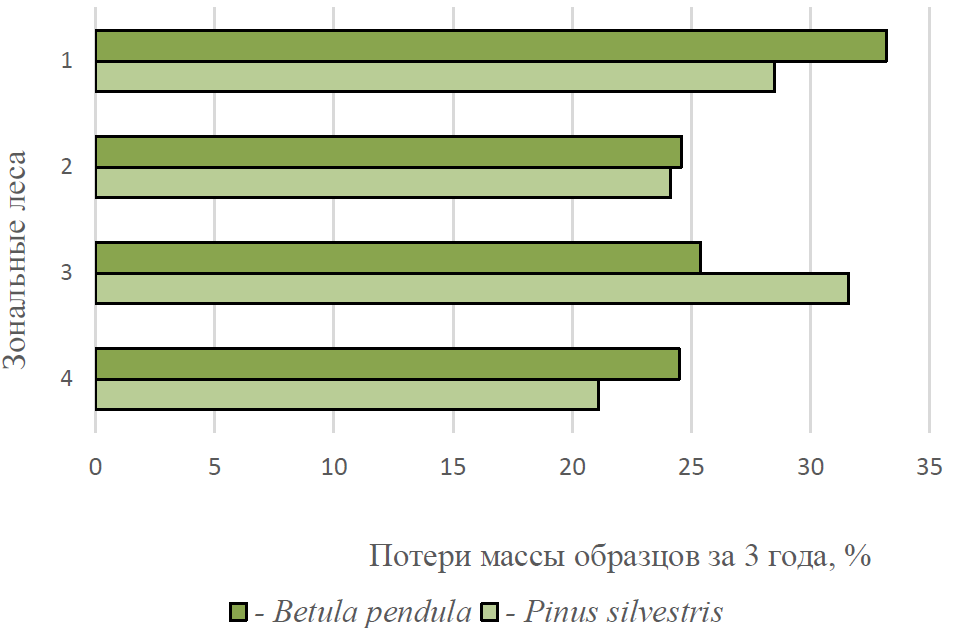
Рис. 3. Скорость микогенной деструкции в зональных лесах
- – южно-таежные темнохвойные леса Западно-Сибирской равнины; 2 – Притобольские лесостепные колки (Мухин, 1993); 3 – леса Южного Предуралья (лесостепная зона) (Сафонов, 2006); 4 – искусственные насаждения Южного Предуралья (степная зона).
Заметное увеличение скорости ксилолиза, отмеченное в Южном Предуралье обусловлено тем, что опыты по изучению скорости разложения древесины сосны были заложены в национальном парке «Бузулукский бор». Его сосновые насаждения являются азональным элементом растительного покрова региона и отличаются специфичными природными условиями.
ЗАКЛЮЧЕНИЕ
Экстраполяция полученных данных о скорости деструкции древесины различных древесных растений в Южном Предуралье на определенный временной промежуток позволяет рассчитать продолжительность процессов деструкции веточного отпада. За конечный уровень деструкции принималась 80 % потеря первоначальной массы древесины (Мухин, 1993). Согласно расчетам, средняя продолжительность процесса утилизации древесины в искусственных насаждениях Южного Предуралья составляет 9 лет.
Таким образом, можно прийти к выводу, что интенсивность микогенного ксилолиза в искусственных насаждениях степной зоны Южного Предуралья относительно высока и вполне сопоставима с таковой в естественных насаждениях. Это свидетельствует о эффективной деятельности системы редуцентов в искусственных насаждениях.
Список литературы
Алексеев В. А. Лесные экосистемы и атмосферное загрязнение. – Л.: Наука. Ленинградское отделение, 1990. – 197 с.
Волчатова И. В., Александрова Г. П., Хамидуллина Е. А. Микогенный ксилолиз в условиях антропогенного загрязнения // Лесоведение. – 2007. – № 5. – С. 27–31.
Диярова Д. К. Углеродно-кислородный газообмен древесного дебриса при микогенном разложении: автороф. дис. … канд. биол. наук: 03.02.08 Экология. – Екатеринбург: Институт экологии растений и животных УрО РАН. – 2020. – 30 с.
Казарцев И. А., Рощин В. И., Соловьев В. А. Разложение углеводов древесины Populus tremula и Picea abies под действием лигнинразрушающих грибов // Микология и фитопатология. – 2014. – Т. 48, № 2. – С. 112–117.
Кузнецов А. А., Капица Е. А. Методика определения запасов и потоков углерода, связанных с крупными древесными остатками в лесах таежной зоны // Известия Санкт-Петербургской лесотехнической академии. – 2009. – № 188. – С. 23–29.
Мухин В. А. Биота ксилотрофных базидиомицетов Западно-Сибирской равнины. – Екатеринбург: Наука, 1993. – 231 с.
Мухин В. А. Дереворазрушающие грибы – современная экологическая парадигма // Биоразнообразие и экология грибов и грибоподобных организмов северной Евразии. – 2015. – С. 170–173.
Сафонов М. А. Скорость микогенной деструкции древесины в лесах Южного Приуралья // Вестник ОГУ. – 2006. – 2 (52). – С. 18–21.
Сафонов М. А., Булгаков Е. А. Вклад микогенной деструкции древесины в формирование микроклимата лесов Южного Приуралья // Фундаментальные исследования. – 2013. – № 10 (12). – С. 2674–2678.
Сафонов М. А., Маленкова А. С., Русаков А. В., Ленева Е. А. Биота искусственных лесов Оренбургского Предуралья. – Оренбург: ООО «Университет», 2013. – 176 с.
Степанова Н. Т., Мухин В. А. Основы экологии дереворазрушающих грибов. – М.: Наука, 1979. – 100 с. Стороженко В. Г. Участие дереворазрушающих грибов в процессах деструкции и формирования лесных
сообществ // Хвойные бореальной зоны. – 2016. – Т. 34, № 1–2. – С. 87–91.
Шорохова Е. В., Капица Е. А., Кузнецов А. А. Микогенный ксилолиз пней и валежа в таежных ельниках // Лесоведение. – 2009. – № 4. – С. 24–33.
Boddy L., Watkinson S. C. Wood decomposition, higher fungi, and their role in nutrient redistribution // Сanadian Journal of Botany. – 1995. – 73, suppl. 1, Sec. E-H. – P. 1377–1383.
Mukhin V. A., Voronin P. Y. Mycogenic decomposition of wood and carbon emission in forest ecosystems // Russian Journal of Ecology. – 2007. – Т. 38. – С. 22–26.
Progar R. A., Schowalter T. D., Freitag C. M., Morrell J. J. Respiration from coarse woody debris as affected by moisture and saprotroph functional diversity in Western Oregon // Oecologia. – 2000. – Т. 124, N 3. – P. 426–431.
Safonov M. A. Mycogenic xylolysis processes in artificial forest plantations in the steppe zone of the Orenburg region (Southern Urals) // Ekosistemy. 2024. Iss. 37. P. 89–95.
Macromycetes are the most significant component of the forest ecosystem decomposers. The process of mycogenic wood decay depends on various factors, which in turn affects the functioning of forest ecosystems. The study of mycogenic xylolysis processes in the Southern Urals (Orenburg region) began in 2000. However, during that period experiment was limited to the forest-steppe part of the region and focused on stands of natural origin. However, the majority of the region is located within the steppe zone, highlighting the relevance of similar studies in the central and southern parts of the region in stands of artificial origin. The experiment was conducted in 2019–2022; so, samples of birch, pine, elm and maple wood were laid for a period of 3 years. The studies show that the rate of mycogenic wood decay varies within the steppe zone of the Southern Urals. The highest rate of decay is characteristic for the branch litter of elm, birch, and pine, while the lowest rate is observed for the decay of maple. A wider variation in the decomposition rate was typical for samples of pine and birch. The decomposition rate of pine wood varies significantly depending on the location, while the indicators for birch wood are more uniform. The comparison of the newly obtained data with previously collected data on the rate of mycogenic xylolysis in the forests of the forest-steppe part of the region indicates that the wood decay rate in artificial plantations is slightly lower than that in natural forests, particularly in relation to pine wood decay. Thus, the intensity of mycogenic xylolysis in artificial plantations of the steppe zone of the Southern Urals is relatively high and is quite comparable to that in natural plantations. This indicates the efficient operation of the system of reducers in artificial plantings.
Key words: macromycete fungi, mycogenic xylolysis, artificial plantations, Southern Urals, Orenburg region.
Поступила в редакцию 05.12.23
Принята к печати 29.02.24
