
Экосистемы, 33: 64–72 (2023) https://ekosystems.cfuv.ru
![]()
УДК 581.41
Державина Н. М.1, Ковынёва С. В.1, Силаева Ж. Г.2
- Орловский государственный университет имени И. С. Тургенева Орел, Россия
d-nm@mail.ru, kovineva_88@inbox.ru
- Орловский государственный аграрный университет имени Н. В. Парахина Орел, Россия
Изучение онтогенеза папоротников вносит существенный вклад в проблемы их биоморфологии, систематики, экологии и физиологии, а также служит основой для популяционнных исследований и выявления закономерностей адаптациогенеза. Целью работы явилось описание онтоморфогенеза спорофита скального папоротника хазмофита Ceterach officinarum. Материалом для исследования послужили сборы, проведённые в Крыму в 2016 году. Применены подходы дискретного описания онтогенеза, разработанные для цветковых растений, с коррективами в терминологии, касающейся специфики репродуктивной биологии папоротников. Охарактеризованы онтогенетические состояния спорофита, начиная с момента появления зародыша после полового процесса и оплодотворения, затем – проростка, ювенильного, имматурного и, заканчивая формированием взрослых структур в надземной и подземной сферах у виргинильного растения. Описан переход папоротника к спороношению у молодых, средневозрастных и стареющих спорофитов, сопровождаемый появлением градатных сорусов на абаксиальной стороне вай. Показан переход к постепенному снижению всех жизненных функций у субсенильного и сенильного спорофитов, ведущий, в конечном счёте, к отмиранию растения. В онтогенезе Ceterach officinarum выявлены следующие фазы морфогенеза: первичной вертикальной розетки у ювенильных растений; неветвящегося розеточного короткокорневищного растения у имматурных и виргинильных спорофитов; ветвящегося розеточного короткокорневищного спорофита, соответствующего возрастным состояниям молодых, зрелых и старых спороносящих растений; короткокорневищной розеточной партикулы, формирующейся в онтогенетических состояниях субсенильных и сенильных растений. Существенная онтогенетическая трансформация формы пластинок вай позволяет говорить о гетеробластии у этого вида.
Ключевые слова: Ceterach officinarum, спорофит, онтоморфогенез, возрастные состояния.
ВВЕДЕНИЕ
Обе фазы гетероморфного жизненного цикла папоротников – спорофиты и гаметофиты исследуются в наши дни как в естественных, так и в лабораторных условиях в аспектах их морфологии, физиологии, экологии, биохимии, популяционной и репродуктивной биологии, эволюции, геносистематики.
Спорофит, как правило, играет доминирующую роль в цикле воспроизведения папоротников, отличаясь от гаметофита не только кормофитным уровнем организации, функционально и габитуально, но и плоидностью, экологией и фитоценотической ролью.
Онтогенез спорофита в случае амфимиксиса, как известно, начинается на гаметофите после процесса оплодотворения, формирования зиготы и зародыша, у которого наблюдается смена типов питания: гетеротрофное уступает место миксотрофному, а затем – автотрофному.
Учитывая то, что в цикле воспроизведения каждая из генераций папоротников проходит собственный онтогенез, его изучение смогло активизироваться лишь после того, как в российской птеридологии применили подходы дискретного описания онтогенеза, разработанные для цветковых растений Т. А. Работновым (1950), А. А. Урановым (1975) и дополненные авторами книг «Ценопопуляции растений …» (Смирнова и др., 1976; Заугольнова и др. 1988).
Подобные исследования папоротников вносят существенный вклад в проблемы их биоморфологии, систематики, экологии и физиологии, а также могут быть основой для
ISSN 2414-4738 Published by V. I. Vernadsky Crimean Federal University, Simferopol
Онтоморфогенез спорофита Ceterach officinarum Willd. (Aspleniaceae)
![]()
популяционнных исследований и выявления закономерностей адаптациогенеза. Однако по сравнению с цветковыми растениями эти сведения продолжают оставаться весьма ограниченными.
- числе первых авторов, применивших концепцию описания онтогенеза спорофитов папоротников, оказались Г. П. Серая с соавторами (1980), Н. М. Державина (1982, 1983), Н. Н. Лащинский и Н. И. Шорина (1985), И. И Гуреева (1990, 2001), Н. М. Державина и Н. И. Шорина (1992), И. Г. Криницын (2004) и др., позднее – и гаметофитов (Шорина, Силантьева, 1998; Гуреева, 2001, 2006; Шорина, 2001; Барабанщикова, 2002, 2007), правда, эти исследования касались преимущественно лесных видов.
Учитывая специфику репродуктивной биологии развивающегося спорофита, птеридологи внесли коррективы в названия его онтогенетических состояний: генеративные растения стали называть «молодыми, зрелыми и старыми спороносящими» (Лащинский,
Шорина, 1985).
- зависимости от морфологического выражения качественных признаков и дополнительных биометрических характеристик, позволяющих идентифицировать отдельные возрастные состояния спорофита, стали использовать: число, форму и размеры его вай, тип филлотаксиса, особенности морфологии ризома, положение в пространстве, тип его ветвления и нарастания, число и расположение кладогенных корней, способность к споровому и вегетативному размножению, соотношение процессов новообразования и отмирания.
- качестве объекта исследования онтоморфогенеза был избран спорофит скального папоротника-хазмофита Ceterach officinarum Willd. – скребницы аптечной (Aspleniaceae), интересной своей толерантностью к понижению увлажнения среды. Это один из немногих обитающих в России пойкилогидричных равноспоровых папоротников (Державина, 2021). Скребница ценна и своим прикладным значением: благодаря богатому химическому составу она широко применяется в народной медицине, а зимостойкость позволяет использовать её в качестве декоративного растения, выращиваемого на альпийских горках (Дикорастущие полезные…, 2001; Коновалова, Шевырева, 2004).
МАТЕРИАЛ И МЕТОДЫ
Материалом для исследования послужили сборы, проведённые в Крыму в Байдарской долине Балаклавского района в окрестностях села Новобобровка на левом берегу реки Бага в 2016 году.
-
- силу того, что у цветковых и папоротников сходны и подчинены единым закономерностям основные этапы индивидуального развития особей, мы воспользовались той же периодизацией онтогенеза с небольшими изменениями (Лащинский, Шорина, 1985; Гуреева, 2001), которая разработана для цветковых растений.
Выделены периоды: пререпродуктивный с онтогенетическими состояниями зародыша (em), проростка (ps), ювенильного (j), имматурного (im) и виргинильного (v) спорофитов; репродуктивный с онтогенетическими состояниями молодого (sp1) средневозрастного (sp2) и стареющего (sp3) спороносящих спорофитов; пострепродуктивный или сенильный с онтогенетическими состояниями субсенильного (ss) и сенильного (s) спорофитов.
При характеристике жизненной формы скребницы использована морфо-экологическая методика И. Г Серебрякова (1962, 1964), Т. И. Серебряковой (1972, 1980), Л. Е. Гатцук (1974)
- синтетическая, объединяющая разные классификационные принципы, предложенная Н. И. Шориной (1994).
Вслед за А. П. Хохряковым (1976, 1979) и Н. И. Шориной (1994) вертикально-розеточными называли папоротники, у которых розетки вай завершают короткие ортотропные ризомы.
Фазы морфогенеза спорофита выделены на основе разработок представителей школы И. Г. Серебрякова.
65
Державина Н. М., Ковынёва С. В., Силаева Ж. Г.
![]()
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Папоротник Ceterach officinarum – средиземноморский петрофитный вид, встречающийся в расселинах скал (хазмофит) на Аравийском полуострове, в Гималаях, в Европе (Атлантической, Центральной, Юго-Восточной (Крым); в Азии: Юго-Западной (Турция, Иран), Средней, Центральной; Средиземноморье, Кавказе. В России: на Северном Кавказе, в Северной Осетии, в Дагестане от уровня моря до субальпики (Красная книга Краснодарского края, 2017).
А. Е. Бобров (1982) относит его к бореально-южноевразиатским видам, встречающимся
- лесном и субальпийском поясах. А. М. Аскеров (1986) включает его на Кавказе в число 19 альпийских папоротников, живущих в трещинах крупных скал и валунов. Встречается не только на скалах, но и на рукотворных стенах. Предпочитает известняки. Оптимум рН лежит
- нейтральной среде, но он может расти как на слабо кислых, так и на слабо щелочных субстратах, то есть является субнейтрофилом бази- и окситолерантным (Gams, 1938). На Северном Кавказе встречается до высоты 3000 м (Литвинская, Муртазалиев, 2013).
- Крыму скребница предпочитает горные местности с климатом от умеренно жаркого полувлажного до прохладного избыточно влажного, обитая в широком диапазоне местообитаний: чаще всего на полном свету, но при этом лучше растёт на речных бортах в
моховых подушках при некотором затенении. Этот папоротник скорее является
теневыносливым растением.
Жизненная форма спорофита – короткокорневищный, радиально-симметричный, вертикально- или косо-розеточный, неявнополицентрический, многолетний, вечнозеленый гемикриптофит (Державина, 2021).
Споры Ceterach officinarum прорастают по Vittaria-типу, а проталлий развивается по Adiantum-типу (Nayar, Kaur, 1971).
Пререпродуктивный период в жизни спорофита начинается с момента появления зародыша спорофита после полового процесса и оплодотворения и завершается полным формированием взрослых структур в надземной и подземной сферах растения.
Зародыш спорофита (em) образуется на гаметофите с момента деления зрелой зиготы, проходит эмбриогенез, образуя на вентральной стороне таллома полярную структуру, как у всех представителей класса Polypodiopsida ориентированную поперечно (Воронин, 1964). Органогенез начинается с формирования зачатка корня, гаустории, зачатка вайи и оси ризома, которые благодаря геотропным движениям приобретают разные направления: будущая вайя
- ось – геометрически вверх, будущий корень – вниз, а гаустория, врастая в ткань гаметофита, обеспечивает гетеротрофное питание зародыша.
Проростки (рs) – миниатюрные розеточные растеньица высотой 3–6 мм связанные с гаметофитом, несущие от 1 до 3–4 длинно-черешковых спирально расположенных вай с пластинками ромбической и округло-почковидной формы (рис. 1). Соотношение длины черешка и пластинки 2:1. Жилка – одна неветвящаяся, либо дихотомирующая. Число придаточных разветвлённых до 2-го порядка корней соответствует числу вай. Черешки и пластинки с обеих сторон покрыты редкими бесцветными чешуйками, которые сохраняются на верхней стороне пластинки вплоть до достижения спороношения. За счёт фотосинтеза и контакта с гаметофитом осуществляется миксотрофное питание. Вайи проростков и спорофитов всех возрастных состояний живут более года, поэтому скребницу можно считать вечнозелёным растением.
Ювенильные (j) растения отличаются несколько бόльшими размерами: 10–20 мм и возросшим до 5–6 числом вай в ортотропной розетке, с учетом образовавшихся у проростков. Форма пластинок вай становится широко-обратно-яйцевидной, соотношение длины черешка
- пластинки 2:1 не изменяется. Чешуи на вайях сохраняются. Порядок ветвления жилок при открытом дихотомическом жилковании возрастает до 3–4. Слабо выраженная терминальная почка ортотропного ризома несёт обычно 1–2 зачатка вай. Корневая система усиливается, так как от основания каждой вайи отходит по корню.
После отмирания гаметофита спорофит переходит к автотрофному питанию.
66
Онтоморфогенез спорофита Ceterach officinarum Willd. (Aspleniaceae)
![]()
Имматурные растения (iт) приобретают некоторые черты взрослых спорофитов, позволяющие определить видовую принадлежность папоротника. При увеличении размеров растения становится заметным ортотропный ризом диаметром 2–3 мм. Отделительный слой между пластинкой и основанием черешка вайи не образуется, поэтому отмирают они постепенно, а в основании ризома сохраняются черешки, сформированные в предыдущих возрастных состояниях. Филлоподии вай входят в состав ризома, обеспечивая его увеличение в диаметре, а черешки прошлогодних вай играют защитную роль для заложенных в верхушечной почке 3–6 зачатков. Число вай составляет 5–6, их длина достигает 20–40 мм. Пластинки приобретают полувзрослые черты, становясь дважды-перистыми. Соотношение длины черешка и пластинки меняется до 1:1, за исключением спорофитов, растущих в густом моховом покрове, у которых черешки вытягиваются, сохраняя соотношение 2:1 и даже 3:1. Порядок ветвления жилок вай достигает 6–7. Корневая система включает все корни (ветвление до 3-го порядка), образовавшиеся за время жизни спорофита.
- взрослых неспороносящих (виргинильных) растений (V) доминируют черты взрослых спорофитов: ортотропный эпигеогенный ризом уже хорошо выражен, его диаметр достигает 3–4 мм и более, он, расширяясь к верхушке, приобретает обратно-узко-конусовидную форму. Емкость терминальной почки возрастает до 5–10 зачатков. Вайи в числе 4–10 на верхушке ризома образуют розетку. Их длина составляет 60–90 мм, пластинки непарноперисторассечённые с 4–5 парами перьев. Перья очередные, но не супротивные, расположены чередуясь по обеим сторонам центральной жилки. У перьев округлая форма, край – слабо волнистый.
Светло-бурые клатратные чешуи, налегающие друг на друга, обильно покрывают черешки, нижнюю поверхность вай и терминальную почку. На верхней стороне сформированных ранее вай они сохраняются только вдоль средней жилки. Чешуи имеют широкое основание, оттянутую верхушку и редкие головчатые клетки по краю (рис. 1). Вайи приобретают жесткость. Соотношение длины черешка и пластинки существенно меняется и составляет 1:2. Тип жилкования максимально приближается к кладодромному, характерному для дефинитивных спорофитов. При этом дихотомирующие жилки разных порядков свободно выходят на край пера.
Ветвление корней достигает двух порядков, формируется первично гоморизная корневая система из кладогенных корней. Участки проксимальной части ризома, возникшие в ювенильном онтогенетическом состоянии, начинают разрушаться или уже разрушились.
Переход особей к спороношению маркирует начало репродуктивного периода Спороносящие растения (sp) отличает появление градатных сорусов на абаксиальной
стороне вай. Сорусы продолговатые, линейные, длиной 3–5 мм, находятся, как и у других видов семейства Aspleniaceae, с одной стороны дихотомирующих жилок. Покрывальце одностороннее линейное, закрывающее весь сорус (рис. 1).
- молодых спороносящих растений (sp1) сорусы располагаются только на верхних перьях вай, у средневозрастных спороносящих (sр2) – на всей пластинке вайи, сорусы находятся среди плотно расположенных чешуй.
Особенностью молодых спороносящих растений является начало ветвления ризома (рис. 1).
Если смотреть на дефинитивные спорофиты сверху, то, в отличие от виргинильных растений, можно обнаружить уже несколько плотно расположенных розеток из 5–(8)–12 вай (рис. 2). Подобная картина создается за счет акрогенного дихотомического ветвления ризома при моноподиальном нарастании осей. Продольные разрезы ризомов позволяют увидеть особенности этого ветвления: на апексе формируются две равноценные дочерние оси, каждая из которых продолжает нарастать моноподиально-дихотомически, при многократном ветвлении возникает расширяющаяся вверх структура, напоминающая «многоярусный дихазий» (Державина, 2021).
67
Державина Н. М., Ковынёва С. В., Силаева Ж. Г.
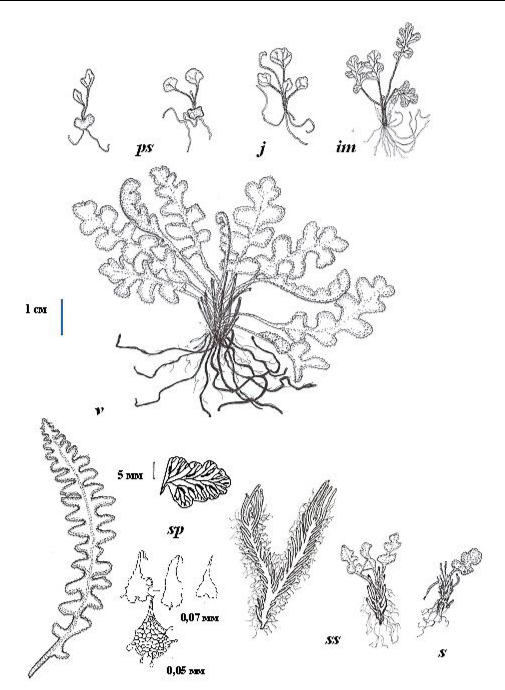
Рис. 1. Онтогенез спорофита Ceterach officinarum
Условные обозначения: рs – проросток; j – ювенильное растение; im – имматурное растение; v – виргинильное растение; sp – спороносящее растение: вайя, перо вайи, чешуи с черешка, фрагмент ризома средневозрастной особи (корни, чешуи и вайи удалены); ss – субсенильное растение; s – сенильное растение.
68
Онтоморфогенез спорофита Ceterach officinarum Willd. (Aspleniaceae)

Рис. 2. Дефинитивный спорофит Ceterach officinarum
Крым, Севастополь, Балаклавский р-н, Байдарская долина, окрестности с. Новобобровка, левый берег реки Бага, май 2016 г. (фото Н. М. Державиной).
Кожистые мономорфные вайи средневозрастных спорофитов зелёного цвета на верхней стороне и бежево-коричневатые отчешуй на нижней. Продолжительность их жизни составляет более года, поэтому этот папоротник можно назвать вечнозелёным.
Основания черешков вай, теряя чешуи, становятся коричневыми. Пластинки достигают максимальных размеров: длина составляет 13–17 см, ширина – 1,5– 2,0 см, число пар перьев варьирует от 10 до 15. Перья имеют округлую, яйцевидную или продолговатую форму, тупую верхушку и лопастной или крупно-волнистый край. Соотношение длины черешка и пластинки – 1:3. Жилки ветвятся до восьмого порядка. Изредка можно наблюдать частичный переход от открытого жилкования к сетчатому, когда появляются ареолы, образованные жилками 6-го порядка. Сорусы, как правило, возникают сбоку от жилок 3-го порядка. Емкость верхушечной почки достигает максимальных значений и составляет 10–12 зачатков вай, которые защищены не только живыми вайями, но и черешками отмерших.
Ветвящиеся до 4–5 порядка корни глубоко внедряются в расселины между камнями. Процессы отмирания на базальном конце ризома продолжаются и наступает момент,
когда ось первого порядка полностью разрушается и происходит партикуляция. Растение при этом не распадается на партикулы, они остаются плотно связанными за счёт переплетающихся корней. Образуется подобие кочки с несколькими верхушками, окруженными розетками вай.
Одной из редких особенностей скребницы является её пойкилогидричность: способность переносить нехватку воды в состоянии криптобиоза. При этом пластинки вай совершают ксеротропные движения, свёртываясь покрытой чешуями нижней поверхностью наружу. В
69
Державина Н. М., Ковынёва С. В., Силаева Ж. Г.
![]()
таком состоянии растение находится до улучшения увлажнения среды, после чего жизненные процессы восстанавливаются.
- стареющих спороносящих растений (sp3) процессы отмирания начинают доминировать над новообразованием. Снижаются интенсивность спороношения и нарастания ризома. Усиливается его разрушение на проксимальном конце, которое ведёт к возрастанию числа укороченных партикул. Растения представляют собой мощные дерновиноподобные кочки с 5–6 и более розетками вай. Емкость терминальной почки, как бы погружённой внутрь розетки, снижается, размеры вай уменьшаются. Форма пластинок остаётся примерно такой же, как и у молодых спороносящих растений. Сорусы образуются только на нескольких верхних перьях вайи.
Наступление постгенеративного периода свидетельствует о переходе к постепенному снижению всех жизненных функций и, в конечном счёте, отмиранию растения.
- субсенильных и сенильных растений (ss, s) наблюдается максимальное упрощение жизненной формы: спороношение отсутствует, а число живых вегетативных структур минимально. Преобладающие процессы разрушения ризома приводят к тому, что субсенильные и сенильные спорофиты представляют собой небольшие чешуйчатые не разветвлённые партикулы или одну коротенькую верхушку оси, увенчанную 2–3 вайями имматурного облика с 2–3 парами перьев. Жилки ветвятся до 2-го порядка. Корневая система состоит в основном из разрушающихся корней, ветвящихся до 2-го порядка.
ЗАКЛЮЧЕНИЕ
Судя по полученным данным, в онтогенезе Ceterach officinarum прослеживаются следующие фазы морфогенеза: первичной вертикальной розетки, формирующейся у спорофитов в ювенильном возрастном состоянии; неветвящегося розеточного короткокорневищного растения, характерного для имматурных и виргинильных растений; ветвящегося розеточного короткокорневищного спорофита, типичного для молодых, зрелых
- старых спороносящих растений; короткокорневищной розеточной партикулы, возникшей у субсенильных и сенильных спорофитов.
Исследование показало, что форма пластинок вай в онтогенезе претерпевает существенную трансформацию вследствие усложнения, а затем – упрощения. Это позволяет говорить о гетеробластии у Ceterach officinarum.
Список литературы
Аскеров А. М. Высокогорные папоротники Кавказа // Растительный покров высокогорий. – Л: Наука, 1986. – С. 6–9.
Барабанщикова Н. С. Онтогенез гаметофитов папоротников и его поливариантность на примере некоторых представителей рода Dryopteris в природных условиях // Труды первой Российской птеридологической конференции и школы-семинара по птеридологии (Томск – Барнаул, 2007). – Томск: Изд-во Томского ун-та, 2007. – С. 12–25.
Барабанщикова. Н. С. Биология спорового размножения некоторых папоротников Подмосковья: автореф.
дис. … канд. биол. наук. 03.00.05: Ботаника. – Москва: МПГУ, 2002. – 16 с.
Бобров А. Е. Папоротникообразные высокогорных экосистем СССР // Пространственная структура экосистем. – Л: Геогр. общ. АН СССР, 1982. – С. 102–109.
Воронин Н. С. Эволюция первичных структур в корнях растений // Ученые записки Калужского государственного педагогического ин-та им. К. Э. Циолковского. – Калуга, 1964. – Вып. 13. – С. 3–179.
Гатцук Л. Е. К методам описания и определения жизненных форм в сезонном климате // Бюллютень МОИП.
Отдел биологический. – 1974. – Т. 79. – Вып. 3. – С. 84–100.
Гуреева И.И. Подходы к изучению онтогенеза равноспоровых папоротников // Материалы VI Всероссийской школы-конференции по водным макрофитам «Гидроботаника 2005» (п. Борок, 11–16 октября 2005 г.). – Рыбинск: ОАО «Рыбинский дом печати», 2006. – С. 87–96.
Гуреева И. И. Равноспоровые папоротники Южной Сибири (систематика, происхождение, биоморфология, популяционная биология). – Томск: Изд-во Томского ун-та, 2001. – 158 с.
Державина Н. М. Онтогенез и структура спорофита Polypodium cambricum L. // Бюллетень МОИП. Отдел биологический. – 1982. – Т. 87. – Вып. 5. – С. 89–101.
70
Онтоморфогенез спорофита Ceterach officinarum Willd. (Aspleniaceae)
![]()
Державина Н. М. Онтоморфогенез спорофита Lepisorus ussuriensis (Regel et Maack) Ching. // Биоморфология растений Дальнего Востока. – Владивосток: ДВНЦ АН СССР, 1983. – С. 63–75.
Державина Н. М. Структурные адаптации устойчивого к десикации папоротника Ceterach officinarum Willd. (Aspleniaceae) Сибирский экологический журнал (Contemporary Problems of Ecology). – 2021. – № 5. – С. 501–511. DOI: 10.15372/SEJ20210501
Державина Н. М., Шорина Н. И. Структура и динамика ценопопуляционных поселений Polypodium vulgare L. в лесах Западного Закавказья // Ботанический журнал – 1992. – Т. 77, № 2. – С.46-54
Дикорастущие полезные растения России. [Ред. А. Л. Буданцев, Е. Е. Лесиовская]. – СПб.: Изд-во СП ХВА,
2001. – 663 c.
Коновалова Т. Ю., Шевырева Н. А. Папоротники для сада. – Москва: Кладезь-Букс, 2004. – 96 с.
Красная книга Краснодарского края. Растения и грибы / [Oтв. ред. С.А. Литвинская и др.]. – Краснодар,
2017. – 850 с.
Криницын И. Г. Онтогенез и структура популяций спорофитов некоторых видов рода Botrychium Sw. в подзонах южной тайги и подтайги Европейской России: автореф. дис. … канд. биол. наук. 03.00.05: Ботаника. – Барнаул, 2004. – 19 с.
Лащинский Н. Н., Шорина Н. И. Онтогенез спорофита и структура ценопопуляций Polystichum braunii (Spenn.) Fee в черневой тайге Салаирского кряжа // Известия СО АН СССР. Серия биологических наук. – 1985. – Вып. 2, № 13. – С. 35–44.
Литвинская С. А., Муртазалиев Р. А. Флора Северного Кавказа: Атлас-определитель. – М.: Фитон XXI,
2013. – 688 с.
Работнов Т. А. Жизненный цикл многолетних травянистых растений в луговых ценозах // Труды БИН АН
СССР. Серия 3: Геоботаника. – 1950. – Вып. 6. – С. 7–204.
Серая Г. П., Комов С. В., Мыльников Н. М., Безденежная Н. Л. Особенности морфогенеза и возрастные состояния спорофита некоторых видов крупнокорневищных лесных папоротников // Онтогенез травянистых поликарпических растений: Межвузовский тематический сборник Уральского ун-та. – Свердловск, 1980. – С. 91– 101.
Серебряков И. Г. Экологическая морфология растений. – Москва: Высшая школа, 1962. – 378 с.
Серебряков И. Г. Жизненные формы высших растений и их изучение // Полевая геоботаника, 1964. – Т. 3. – С. 146–205.
Серебрякова Т. И. Учение о жизненных формах растений на современном этапе // Итоги науки и техники. – Москва: ВИНИТИ. – Серия «Ботаника», 1972. – Т. 1. – С.84–169.
Серебрякова Т. И. Еще раз о понятии «жизненная форма» у растений // Бюллетень МОИП. Отдел биологический. – 1980. – Т. 85. – Вып. 6. – С. 75–86.
Уранов А.А. Возрастной спектр ценопопуляций как функция времени и энергетических волновых процессов // Научные доклады высшей школы «Биологические науки». – 1975. – № 2. – С. 7–34.
Хохряков А. П. Жизненные формы папоротникообразных и возможные пути их эволюции // Материалы V Московского совещания по филогении растений – Москва: Наука, 1976. – С. 184–187.
Хохряков А. П. Жизненные формы папоротникообразных, их происхождение и эволюция // Известия АН СССР. Серия биологическая. – 1979. – № 2. – С. – 251–264.
Смирнова О. В., Заугольнова Л. Б., Ермакова И. М. и др. Ценопопуляции растений (основные понятия и структура) – М.: «Наука», 1976. – 216 с.
Заугольнова Л. Б., Жукова Л. А., Комаров А. С. и др. Ценопопуляции растений: (очерки популяционной
биологии). – М.: «Наука», 1988. – 184 с.
Шорина Н. И. Экологическая морфология и популяционная биология представителей подкласса Polypodiidae: автореф. дис. … д-ра биол. наук. 03.00.05: Ботаника.– Москва: МГУ, 1994. – 34 с.
Шорина Н. И. Популяционная биология гаметофитов равноспоровых папоротников Polypodiophyta // Экология. – 2001. – № 3. – С. 182–187.
Шорина Н. И., Силантьева Л. А. Особенности онтогенеза некоторых представителей рода Asplenium (Aspleniaceae) в связи с их ксерофилизацией // Проблемы ботаники на рубеже ХХ–ХХI веков: тез. докл., представленных на II (Х) съезде Русского ботанического общества. – Санкт-Петербург, 1998. – Т. 1. – С. 93.
Gams S. H. Ökologie der extratropischen Pteridophyten / [Ed. F. Verdoorn)]. – Manual of Pteridology. – M. Nijhoff, The Hague, 1938. – P. 382–419.
Nayar B. K., Kaur S. Gametophytes of gomosporous ferns // Botaical Rewiew. – 1971. – Vol. 37, N 3. P. – 295–396.
71
Державина Н. М., Ковынёва С. В., Силаева Ж. Г.
![]()
Derzhavina N. M., Kovyneva S. V., Silaeva Zh. G. Ontomorphogenesis of the sporophyte Ceterach officinarum Willd. (Aspleniaceae) // Ekosistemy. 2023. Iss. 33. P. 64–72.
The study of ferns ontogenesis makes a significant contribution to solution of issues of their biomorphology, systematics, ecology and physiology, and also it serves as the basis for population studies and identification of patterns of adaptatiogenesis. The aim of this research is to describe ontomorphogenesis of the sporophyte of the rock fern Ceterach officinarum. The material for the study was collected in Crimea in 2016. The approach of discrete description of ontogenesis developed for flowering plants was applied for the research but some adjustments were made in terminology concerning the specifics of the reproductive biology of ferns. The age-related conditions of the sporophyte were described starting from the moment of an embryo appearance after sexual process and fertilization, then – the seedling, juvenile, immature, and ending with the complete formation of adult structures in the aboveground and underground spheres of the virginal plant. The transition of ferns to sporulation in young, middle-aged and aging sporophytes, accompanied by appearance of graded soruses on the abaxial side of fronds is characterized. Moreover, the paper outlined transition to a gradual decrease in all vital functions in the subsenile and senile sporophytes, leading, ultimately, to the death of the plant. The following phases of morphogenesis were revealed in the ontogenesis of Ceterach officinarum: primary vertical rosette in juvenile plants; non-branching rosette of short-rhizomatous plants in immature and virginal sporophytes; branching rosette of short-rhizomatous sporophyte corresponding to the age conditions of young, mature and old spore-bearing plants; short-rhizomatous rosette particula formed in ontogenetic conditions of subsenile and senile plants. A significant ontogenetic transformation of the shape of the fronds plates indicates heteroblasty in this species.
Key words: Ceterach officinarum, sporophyte, ontomorphogenesis, age-related conditions.
Поступила в редакцию 05.03.23
Принята к печати 20.03.23
72
