
МОРФОМЕТРИЧЕСКИЕ ИССЛЕДОВАНИЯ КУЛЬТУРЫ ARTHROSPIRA РLATENSIS ПРИ ДЕФИЦИТЕ МИНЕРАЛЬНОГО ПИТАНИЯ
MORPHOMETRIC STUDIES OF ARTHROSPIRA PLATENSIS CULTURE AT DEFICIENCY OF MINERAL NUTRITION
JOURNAL: «EKOSISTEMY», Issue 40, 2024
Publication text (PDF): Download
UDK: 579.2:582.232
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Kharchuk I. A., Rylkova O. A.
A. O. Kovalevsky Institute of Biology of Southern Seas, Russian Academy of Sciences, Sevastopol, Russia
TYPE: Article
DOI: https://doi.org/10.29039/2413-1733-2024-40-7-13
PAGES: from 7 to 13
STATUS: Published
LANGUAGE: Russian
KEYWORDS: cyanobacteria, Arthrospira (Spirulina) platensis, trichomes, adaptation, cyanobacterial biomass.
ABSTRACT (ENGLISH):
Lack of mineral nutrition belongs to the main natural stress factors and plays an important role in the life activity of aquatic organisms. In the cultivation of microalgae and phototrophic prokaryotes, organisms are forced to develop various adaptive mechanisms when the environment is depleted as a result of nutrient consumption to maintain their vital activity. Standard methods of cyanobacteria cultivation, optical and microscopic approaches were used in the study of Arthrospira (Spirulina) platensis (Nordstedt) Gomont (1892). The aim of this research was to examine morphometric characteristics of cyanobacteria during long-term accumulative cultivation under conditions of mineral nutrition deficiency. It was experimentally revealed that the density of A. platensis culture during the experiment depended on the external conditions: temperature (T) and illumination (Ilm). During the period of 1–28 days, the density of the culture increased gradually to 0.11 g·l-1 at minimum temperature and illumination (variant 1A); the curve of biomass accumulation had an S-shaped form, an increase to 0.69 g·l-1 and 1.3 g·l-1 was observed at increasing and maximum values of T and Ilm (variants 2A and 3A). After A. platensis culture entered the stage of inactive growth (29–85 days), the biomass accumulation trend continued: in variant 1A, the gradual increase of cyanobacteria density up to 0.42 g·l-1 continued; in variants 2A and 3A, the biomass increased unevenly and reached 2.0 g·l-1 and 5.5 g·l-1, respectively. By the end of the experiment, the colour of culture changed from emerald to dark green, green-brown and yellow-brown. It was revealed that between days 36 and 85 of the experiment in variant 1A, trichomes with the size of 50–100 µm (44.4–61.3 %) dominated more often, the share of cells with minimal filament length (less than 50 µm) was 30–53.3 %. At increasing T and Ilm during the periods of cyanobacteria biomass growth, small trichomes dominated (43.4–85.1 %), but larger filaments over 100 µm also appeared (2.1–22 %); at stationary stages, the share of small cells was also high (51–93 %), large trichomes were not found or their contribution was halved. This trend is consistent with the data described earlier. The most pronounced changes in the contributions of morphological groups were observed at maximum T and Ilm. Probably, the lack of biogens triggered adaptation mechanisms of cyanobacteria – trichome fragmentation.
ВВЕДЕНИЕ
Факторы среды имеют минимальные и максимальные значения, за пределами которых организм не может существовать. Недостаток минерального питания относится к основным природным стрессовым факторам, который связан, прежде всего, с дефицитом азота, серы или фосфора, входящих в состав аминокислот, фотосинтетических пигментов, нуклеиновых кислот, коферментов, витаминов и других соединений (Гапочка, 1991). Минеральный стресс играет важную роль в жизнедеятельности фитопланктонных организмов, значительная часть которых обитает в обедненных биогенными элементами районах Мирового океана. Известно, что при культивировании микроводорослей и фототрофных прокариот, основными факторами для их роста и развития являются температура, освещенность и оптимальное насыщение культуральной среды биогенами. При обеднении среды, в результате расхода биогенов организмы вынуждены адаптироваться, чтобы поддерживать свою жизнедеятельность (Шахматов, Ефимова, 2005; Харчук, Алисиевич, 2010). В настоящий момент ощущается необходимость системных исследований структурно-функциональных изменений микроводорослей и цианобактерий в условиях дефицита минерального питания для выяснения основных стратегий, которые используются ими для адаптации к стрессу. Исследований, посвящённых изучению адаптационных свойств Arthrospira рlatensis, много (Van Eykelnburg, 1979; Кондратьева, 1989; Жарикова и др., 2018), однако лишь незначительное количество работ посвящено динамике размерного состава цианопрокариот (Шахматов, Ефимова, 2005; Харчук, Алисиевич, 2010).
Целью данной работы стало исследование динамики морфометрических характеристик A. рlatensis на стадии неактивного роста в условиях дефицита минерального питания.
МАТЕРИАЛЫ И МЕТОДЫ
Растительный материал, Объектом исследования служила альгологически чистая неаксеничная культура Arthrospira (Spirulina) platensis (Nordstedt) Gomont (штамм IBBS–31) из коллекции отдела биотехнологии и фиторесурсов ФИЦ ИнБЮМ. Музейную культуру А. platensis концентрировали путем фильтрации через мельничный газ 100–105 ПЭ, затем трижды промывали дистиллированной водой (в соотношении 1:3) и дважды питательной средой. Для дальнейшего очищения пасту А. platensis помещали в пробирки с питательной средой и центрифугировали 5 мин при 3000 об.·мин-1 (центрифуга ОПН–3-УХЛ 42, Россия), супернатант удаляли, к осадку вновь добавляли питательный раствор, процедуру повторяли трижды. Далее пасту А. platensis (2 мл) переносили в стеклянные конические колбы объёмом 0,5 дм3, объём среды составлял 0,3 дм3 при высоте слоя 6 см. Условия культивирования цианобактерий представлены в таблице 1, во всех вариантах для культивирования А. platensis использовали среду Заррук (Faucher et al., 1979).
Таблица 1
Условия культивирования Arthrospira platensis
| № пробы | Температура | Освещенность |
| 1 А | 14–16 ºС | 0,2 кЛк |
| 2 А | 20–22 ºС | 1,1 кЛк |
| 3 А | 25–27 ºС | 10 кЛк |
Определение биомассы цианобактерий. Содержание сухого вещества (СВ) определяли расчетным методом по уравнению регрессии. Для перехода от единиц оптической плотности (D750) к величине сухого вещества, использовали эмпирический коэффициент для А. platensis (Геворгиз и др., 2005).
Морфологические исследования. Микроскопические исследования трихом А. platensis осуществляли с помощью светового микроскопа Carl Zeiss («Axiostar plus», Германия), снабженного камерой («Сanon А 620», Япония) при увеличении от ×200 до ×630. Размеры трихом А. platensis определяли с помощью программы Image J 1.50i program (National Institutes of Health, United States, Java 1.6.0_20 (32-bit)). Всего измерено 2400 трихом, рассчитан вклад каждой из размерных групп в общую численность.
Статистический анализ проводили в пакете STATISTICA (data analysis software system), версия 10 («StatSoft. Inc.», www.statsoft.com), построение графиков – в программах SigmaPlot 10,0 («SYSTAT Software. Inc.») и Grapher 8 («Golden Software, Inc.»).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ
В течение первых 28 суток эксперимента при минимальных температуре и освещенности (вариант 1А), прирост биомассы был постепенным и минимальным, плотность А. platensis составляла 0,11 г·л-1. При повышении и максимальных Т и Осв (2А и 3А, соответственно) накопительная кривая имела S-образный вид, включающий: лаг-, лог-фазы, линейный рост, период замедления, стационарную фазу. Плотность биомассы в конце стационарной фазы роста для 2А составляла 0,69 г·л-1, для 3А – 1,3 г·л-1, что в 6,3 и 11,8 раза выше, чем в варианте 1А. После выхода на стационарную фазу, были продолжены наблюдения за культурами А. platensis уже вне стадии активного роста (29–85 суток). В варианте 1А биомасса культуры продолжала плавно нарастать (до 0,42 г·л-1); при повышении температуры и освещенности были выявлены скачкообразные периоды повышения плотности культуры цианобактерий: в варианте 2А к концу эксперимента биомасса составляла 2,0 г·л-1, в варианте 3А – 5,5 г·л-1, то есть в 4,8 и 13,1 раз больше, чем при минимальных значениях Т и Осв.
Аналогичные кривые роста для штаммов Arthrospira рlatensis из коллекции ФИЦ ИнБЮМ (штаммы IBSS-31, IBSS-32; накопительное культивирование; Т=28–32 ºС, Осв=8 кЛк) были описаны ранее (Шахматов, Ефимова, 2005). Авторами показано, что до 27 суток кривая роста имела стандартный S-образный вид, максимальная биомасса (около 1,3 г·л-1) была обнаружена на 45–53 сутки, далее (до 60 суток), происходило небольшое снижение плотности культуры цианобактерий до биомассы менее 1 г·л-1. Меньшие абсолютные величины, по сравнению с нашими данными, вероятно, были связанны с большим объемом культиватора (6 л) и, соответственно, неполным исчерпанием биогенных элементов в среде.
Размерная структура культуры А. platensis. Начиная с 36 по 85 сутки исследований, нами проведены морфологические исследования суспензии А. platensis. Установлено, что в культуре А. platensis встречались нити различных размеров (рис. 1 a – c), их диапазон изменялся от 4 мкм до 491 мкм, кроме того, визуализировались фрагменты разрушенных трихом, их максимальное количество обнаружено в варианте при максимальных Т и Осв.
Морфометрический анализ показал, что соотношение трихом различной длины в динамике роста было неодинаково. Так, при минимальной температуре и освещенности (1А) на протяжении всего эксперимента преобладали трихомы размером 0–50 мкм (33–54 %) и 50–100 мкм (44–63 %). Вклад трихом размером 100–150 мкм был незначителен и составлял 2–13 %. С 55 суток культивирования отмечено появление нитей размером 150–200 мкм (1–1,5 %) (рис. 2 a).
В культуре, выращиваемой при повышенной температуре и освещенности (2А), на 36 день эксперимента преобладали мелкие трихомы (0–50 мкм), составляя 50 % от общего числа нитей, к 41 суткам отмечено увеличение их доли до 90 %, вероятно, за счёт деструкции более крупных нитей 50–100 мкм. В этот период обнаружены три размерные группы – кроме мелких (85,2 %) и средних нитей (12,8 %) появились более крупные трихомы размером 100–150 мкм (2,1 %). Далее, к 48 – 62 суткам, также доминировали трихомы размером 0–50 мкм, отмечен количественный рост филаментов 50–100 мкм до 10–30 %, нити размером 100–150 мкм не зарегистрированы. На 70 сутки, на фоне снижение плотности культуры, зафиксировано увеличение доли трихом размером 0–50 мкм до 88,5 %. В период 77 – 85 суток выделяли только нити размером 0–50 мкм и 50–100 мкм, их соотношение варьировало 72–83 % и 28–17 %, соответственно (рис. 2 b).
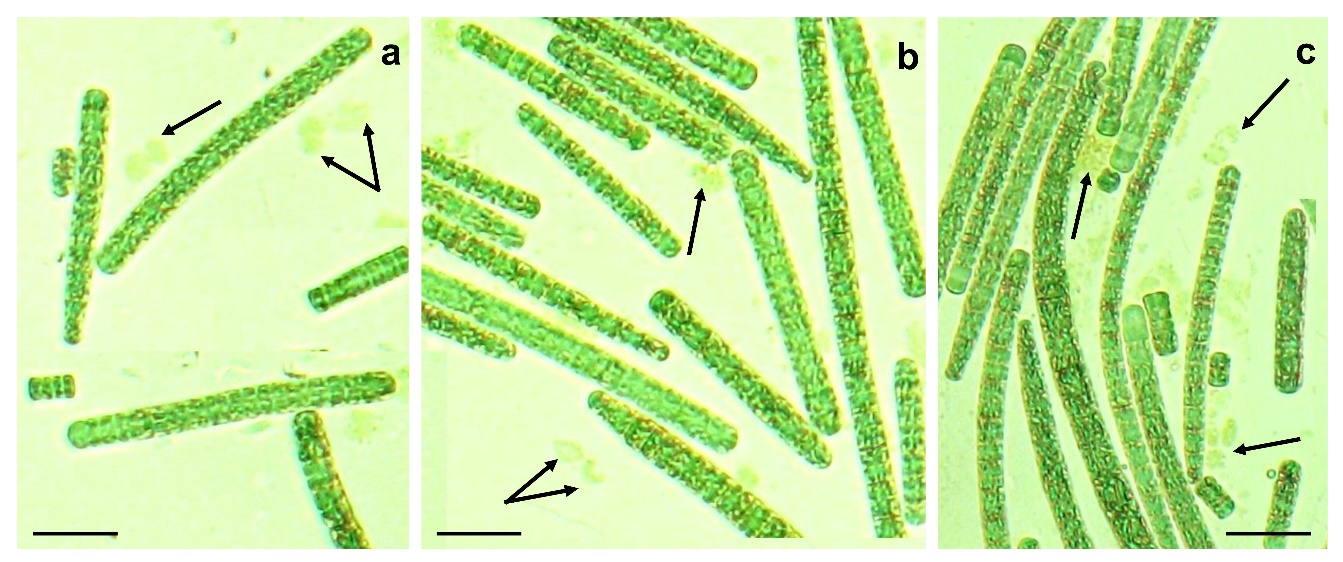
Рис. 1. Общий вид трихом в культуре Arthrospira platensis на 55 сутки эксперимента
Варианты: 1А (a); 2А (b); 3А (с). Стрелками указаны фрагменты разрушенных трихом. Масштабный отрезок – 10 мкм.
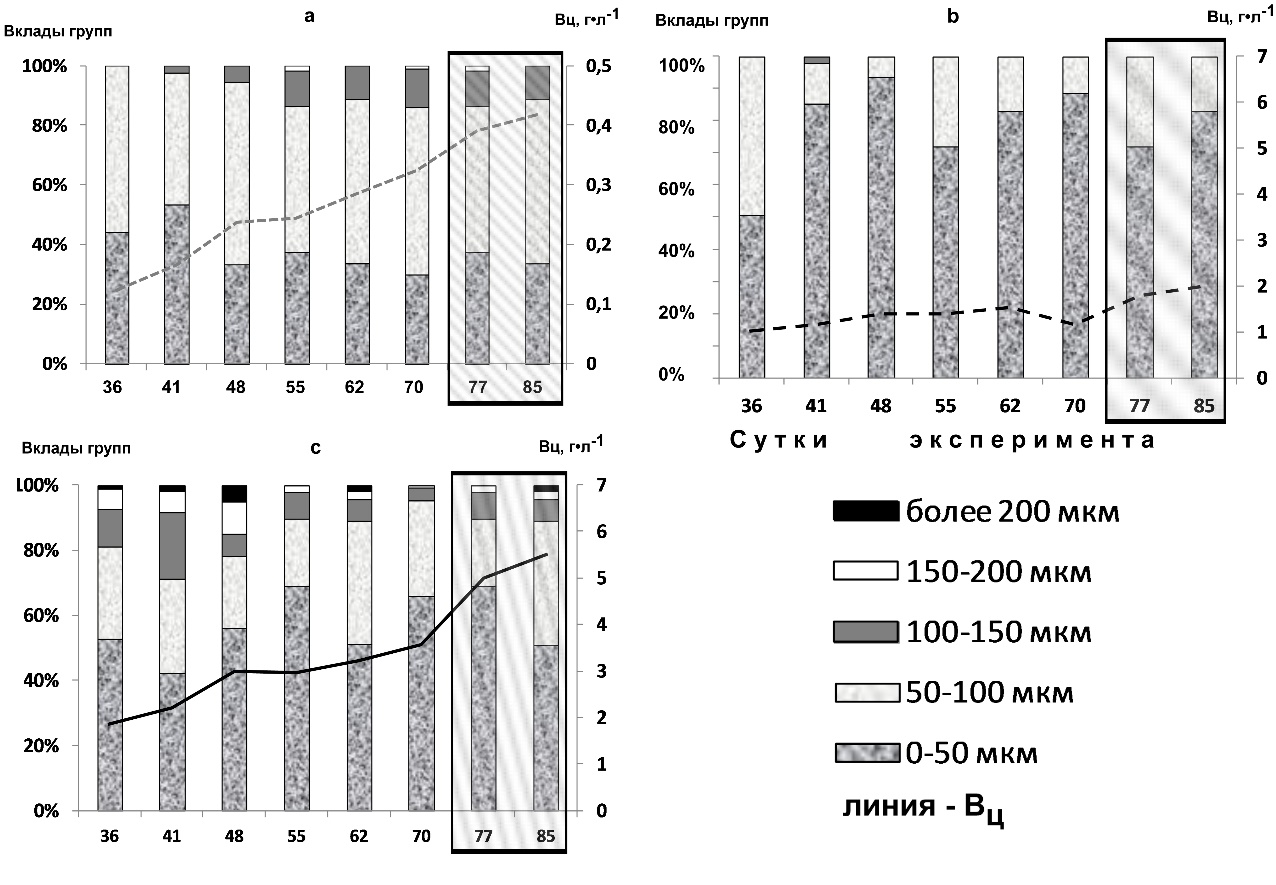
Рис. 2. Изменения биомассы цианобактерий (Вц) и распределение вкладов трихом различной длины в культуре Arthrospira platensis на протяжении 36–85 суток исследований
Варианты: 1А (a); 2А (b); 3А (с). Заштрихованная область – изменение цвета культуральной суспензии.
При максимальных Т и Осв (3А), на 36 – 41 сутки трихомы были представлены всеми размерными группами: 0–50 мкм: 50–100 мкм : 100–150 мкм : 150–200 мкм : 200 мкм и более, их соотношение варьировало в пределах 42,4–56 % : 22–28,8 % : 7–20,3 % : 6,3–10 % : 1–5 %. Характерно, что в период 48 – 62 суток (при стабилизации биомассы) долевое соотношение нитей размером более 100 мкм снижалось в 1,8–3 раза. При увеличении плотности культуры (70 – 77 суток) преобладали мелкие нити размером 0-50 мкм (64–68%) и 50–100 мкм (21–29 %), выявлены нити размером 100–150 мкм и 150–200 мкм, хотя их вклад был незначителен и составлял 4–8 % и 0,7–2% соответственно. К 85 суткам эксперимента отмечено появление нитей длиной более 200 мкм (1,8 %). Показатели для групп 0–50 мкм и 100–150 мкм снижались в 1,3 раза, а в группе размером 50–100 мкм и 150–200 мкм – увеличивались в 1,8 и 1,3 раза соответственно (рис. 2 c).
В целом, для варианта 1А зафиксировано доминирование трихом размером 50–100 мкм (53,6±4,7 %), тогда как при повышении Т и Осв максимальным был вклад самой мелкой фракции (до 50 мкм): 84,5±6,4 % и 55,8±7,6 %, соответственно для 2А и 3А.
Известно, что ультраструктура и морфология цианобактерий сильно зависят от условий среды и факторов питания. С повышением температуры культивирования увеличивались высота и диаметр спирали трихома, а оболочка, окружающая нить, начинала лучше обозначаться. Повышение освещенности способствовало удлинению трихом и фикобилисом (Van Eykelnburg, 197; Жарикова и др., 2018). При исследовании размерных характеристик трихом различных штаммов Arthrospira рlatensis, начиная со стадии адаптации до стационарной фазы роста (около 30 суток), выявлена тенденция возрастания доли крупных трихом на линейной стадии роста и стабилизация показателей на стационарной фазе культивировании (Шахматов, Ефимова, 2005: Жарикова и др., 2018). Единичные исследования по изучению процессов роста после активной фазы накопления биомассы цианопрокариотов (Шахматов, Ефимова, 2005) показали, что во время культивирования Arthrospira рlatensis IBSS-32 в течение 50–60 суток доминировали мелкие трихомы, на фазе отмирания доля коротких трихом увеличивалась за счёт снижения вклада длинных и средних нитей. Изменение морфологических характеристик цианобактерий на поздних этапах культивирования авторы связывали с исчерпанием биогенов из культуральной среды. Известно, что при полном исключении из питательной среды магния и азота происходила фрагментация трихом и уменьшение ширины оборота и длины спирали (Кондратьева, 1989; Жарикова и др., 2018).
В нашем исследовании (штамм IBSS-31), начиная с 36 суток, при минимальных Т и Осв весь период доминировали трихомы размером 50–100 мкм (49,3–61,3 %), короткие нити менее 50 мкм (30–53,3 %). А образцах, выращиваемых при повышенных Т и Осв (2А и 3А), после стадии активного роста цианобактерий доминировали мелкие трихомы (43,4–85,1 %), но появились и более крупные нити длиннее 100 мкм (2,1–22 %); при стабилизации абсолютных величин плотности культуры Arthrospira рlatensis доля мелких клеток также была высока (51–93 %), крупных трихом обнаружено не было или их вклад снижался вдвое, что согласуется с данными других исследователей (Шахматов, Ефимова, 2005: Жарикова и др., 2018).
Необходимо отметить, что во всех вариантах эксперимента в последние две недели мы наблюдали изменение цвета культуральной суспензии с изумрудного на темно-зеленый и зелено-коричневый (для 1А, 2А, соответственно); и желто-коричневый (3А) (рис. 2, заштрихованные области). Это согласуется с аналогичными исследованиями, когда при длительном культивировании A. platensis в исходном питательном растворе без пересева при истощении запасов биогенов наблюдали изменение окраски в сторону красноватых оттенков (Сиренко, Третьяков, 2005).
Измельчение трихом, полученное в нашем исследовании, вероятно, также было связано с недостатком биогенов, так как основная их часть была израсходована цианопрокариотами на поддержание своей жизнедеятельности (Харчук, Алисиевич, 2005). Такая фрагментация нитей, по-видимому, являлась реакцией культуры цианобактерий на недостаток минерального питания. Аналогичная тенденция дробления трихом А. platensis была описана и при других стрессовых условиях. Например, при добавлении в культуру цианобактерий, находящихся на длительной стационарной фазе роста, свежего питательного раствора, содержащего высокие концентрации биогенов, провоцировало их осмотический шок. Подобная фрагментация нитей цианобактерий происходила и в «старых» культурах, культивируемых более 30 суток и без добавки питательной среды (Харчук, Алисиевич, 2005). Вероятно, стрессовые ситуации, вызванные различными внешними факторами (недостаток или, наоборот, высокие концентрации биогенов), запускали однотипные механизмы адаптации – фрагментацию трихом.
Известно, что при небольших скоростях роста альгокультур обычно отсутствует эффект «затенения», и в среде присутствует достаточное количество биогенов (Тренкеншу, Лелеков, 2017). Это обнаружено нами при минимальном (но достаточном для осуществления процессов фотосинтеза) освещении и низкой температуре (1А), когда трихомы не ограничены в питании, делятся неинтенсивно, растут замедленно и достигают размеров более 150–200 мкм. При более благоприятных условиях для роста (варианты 2А, 3А), плотность культуры нарастает быстрее, в определённое время возникает лимит по освещению и биогенам (Тренкеншу, Лелеков, 2017), культура выходит на стационарную фазу роста и, вероятно, запускаются процессы адаптации. Трихомы разрываются по специальным некридиальным клеткам (Vonshak, A., Ed., 1997), образуя маленькие филаменты, для жизнедеятельности которых условия освещенности и температуры оптимальны.
ЗАКЛЮЧЕНИЕ
Экспериментально установлено, что плотность культур A. рlatensis при длительном накопительном культивировании (85 суток) зависела от внешних условий: температуры и освещенности. Выращивание A. рlatensis (1А) при температуре и освещенности ниже оптимума, плотность культуры в период 1–28 суток плавно возрастала до 0,11 г·л-1, при повышении Т и Осв до оптимальных и выше (варианты 2А и 3А) кривая накопления биомассы имела S-образный вид, наблюдали увеличение до 0,69 г·л-1 и 1,3 г·л-1. После выхода культуры A. рlatensis на стадию неактивного роста (29–85 сутки) сохранялась тенденция накопления биомассы: в варианте 1А продолжалось постепенное увеличение плотности цианобактерий до 0,42 г·л-1; в вариантах 2А и 3А биомасса увеличивалась неравномерно и достигала
2,0 г·л-1 и 5,5 г·л-1. К концу эксперимента зафиксировано изменение цвета культуры A. рlatensis, наиболее ярко выраженное в варианте, выращиваемом при максимальных Т и Осв. Морфометрический анализ филаментов показал, что на стадии неактивного роста, независимо от условий выращивания, в культурах преобладали мелкие трихомы размером 0–50 мкм (37,8–84,5 %) и 50–100 мкм (37,8–84,5 %). Доля трихом длиной 100–150 мкм варьировала в пределах 0,42–9,6 %. Трихомы размером 150–200 мкм и более 200 мкм были малочисленны: в 1А их доля составляла 0,5 %; в 3А – 4,8 % и 1,6%. В период возрастания оптической плотности культуры после завершения стандартного накопительного культивирования A. рlatensis обнаружено доминирование мелких, реже средних трихом, а также увеличение доли более крупных нитей; при стабилизации абсолютных величин плотности культуры Arthrospira рlatensis зафиксировано снижение вклада более крупных трихом. Вероятно, стрессовая ситуация, вызванная недостатком биогенов, запускала механизм адаптации цианопрокариот – фрагментацию трихом.
Работа выполнена в рамках государственного задания ФИЦ ИнБЮМ по теме «Комплексное исследование экологических и физиолого-биохимических особенностей микроводорослей различных таксономических групп при адаптации к меняющимся условиям среды» (№ гос. регистрации 124021300070-2).
- Faucher O., Coupal В., Leduy A. Utilization of scawater – urea as a culture medium for Spirulina maxima // Canadian Journal of Microbiology– 1979. – Vol. 25. – P. 752–759.
- Гапочка Л. Д. Об адаптации водорослей. – М.: Изд. Моск. ун., 1981. – 80 с.
- Геворгиз Р. Г., Алисеевич А. В., Шматок М. Г. Оценка биомассы Spirulina platensis (Nordst.) Geitl по оптической плотности культуры // Экология моря. – 2005. – № 70. – С. 96–106.
- Жарикова Г. Г., Елисеева Л. Г., Рыжакова А. В., Ширшов А. Т., Скрипникова М. И., Юрина О. В., Леонова И. Б. Инновационные белковые продукты на основе спирулины – решение продовольственной проблемы. – М.: КноРус, 2018. – 218 с.
- Харчук И. А., Алисиевич А. В. Адаптация Spirulina platensis к изменениям культуральной среды // Экология моря. – 2010. – Т. 80. – С. 92–99.
- Шахматов А. П., Ефимова Т. В. Динамика размерного состава нитчатой цианобактерии Spirulina platensis в накопительной культуре // Экология моря. – 2005. – Т. 70. – С. 84–89.
- Кондратьева Н. В. Морфология популяций прокариотических водорослей. – Киев: Наук. думка 1989. – 175 с.
- Кондратьева Н. В. Строение протопласта клеток Cyanophyta (Обзор литературных данных) // Альгология. – 1994. – Т. 4, № 2. – С. 84–98.
- Сиренко Л. А., Третьяков О. Л. Спирулина и ее использование в биотехнологии // Экология моря. – 2005. – Т. 70. – С. 42–48.
- Тренкеншу Р. П., Лелеков А.С. Моделирование роста микроводорослей в культуре. – Белгород: ООО «КОНСТАНТА», 2017. – 152 с.
- Van Eykelenburg, C. The ultrastructure of Spirula platensis in relation to temperature and light intensity. // Antonie Van Leeuwenhoek. – 1979. – V. 45. – P. 369–390.
- Vonshak A. Spirulina: Growth, Physiology and Biochemistry // Spirulina platensis (Arthrospira): Physiology, Cell-biology and Biotechnology. – London: Taylor & Francis, 1997. – P. 43–65.
