
МОНИТОРИНГОВЫЕ ИССЛЕДОВАНИЯ В ОПУКСКОМ ЗАПОВЕДНИКЕ. АНТЭКОЛОГИЧЕСКИЙ АСПЕКТ
MONITORING STUDIES IN THE OPUKSKY NATURE RESERVE. THE ANTECOLOGICAL ASPECT
JOURNAL: «EKOSISTEMY», Issue 39, 2024
Publication text (PDF): Download
UDK: 574.4:581.162.3(477.75)
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Abramenkov A. A.1, Korzhenevsky V. V.1, Ivanov S. P.2, Korzhenevskaya Yu. V.1
1Federal State Budgetary Institute of Science «The Order of the Red Banner of Labour Nikita Botanical Gardens — National Scientific Center RAS», Yalta, Russia
2V.I. Vernadsky Crimean Federal University, Simferopol, Russia
TYPE: Article
DOI: https://doi.org/10.29039/2413-1733-2024-39-73-83
PAGES: from 73 to 83
STATUS: Published
LANGUAGE: Russian
KEYWORDS: monitoring, pollination of plants, Opuksky Nature Reserve.
ABSTRACT (ENGLISH):
A comparative characteristic of the model profile laid on Mount Opuk in 1988 (the year the profile was laid) and in 2023 is presented. The locations of 40 geobotanical descriptions, ranging from sea level to the northwestern slope, are indicated. The numerical composition of groups of plant species differentiated by the method of pollination is assessed, and divided into four main types: entomogamy, anemogamy, autogamy and hydrogamy, each associated with a varying number of pollination vectors. Changes in the ratio of vectors and pollination methods for more than 200 plant species were evaluated at the profile sites. The assessment of the ecotope conditions at the description sites was carried out according to a specially developed program, with information components sourced from the Ecodata database. It was found that for the period from 1988 to 2023 with a total length of 34 years, the conditions on the gradients of the environmental factors of the edaphotope did not change, the climate did not change as a whole, but small deviations were specified.Significant changes were recorded in the number and structure of species groups of different pollination methods. There was a ten percent or more shift in the ratio of entomogamy and anemogamy towards the latter with more or less stable autogamy. It is suggested that the revealed changes in the ratio of entomogamous and anemogamous species occurred as a result of a significant decrease in the density of pollinators, in particular, wild solitary bees, recorded in the steppe and foothills areas of Crimea in recent decades. The decrease in bee density is due to air saturation with microplastic particles clogging the respiratory tract of insects. The dynamics of changes in the composition of the phytocenosis, revealed through the analysis of the structure of the phytocenosis in relation to pollination methods, indicates the need for further monitoring of the transformation processes of the reserve ecosystem. The results of the study help to understand what changes occur in plant communities under the influence of various factors, including climate change and anthropogenic impact. The research emphasizes the importance of long-term monitoring of changes in natural communities and reveals what methods and approaches can be used to study these changes.
ВВЕДЕНИЕ
Антэкология, как отмечает А. Н. Пономарев (1969), изучает связь между цветущим растением и средой обитания и вскрывает зависимость опыления не только от агентов, осуществляющих его, но и от других, действующих косвенно экологических факторов. Экосистемы трансформируются во времени (на хронотренде) и совокупность растений выступает индикатором их изменений под действием абиотических и биотических факторов (Иванов, 1995).
Поскольку особи растений, так же, как и их совокупности (популяции, сообщества), приурочены к определённым местообитаниям, организованным градиентами факторов среды и представляющими собой некий гиперобъем, то можно считать, что границы толерантности видов могут попадать при изменениях условий, как в зону оптимума, так и в зону пессимума (Плугатарь, Корженевский, 2017). При этом отдельные виды будут либо внедряться в ценоз, либо элиминироваться из него. Экологическая ниша (место в ценозе) у разных видов может быть узкой и широкой. Её размеры положены в основу различения видов растений – стенотопов и эвритопов. Стенотопы обладают меньшей пластичностью, низким уровнем генетического полиморфизма и поэтому более чувствительны к изменению параметров среды. При незначительных трансформациях факторов среды они попадают в зону дискомфорта, в то время как эвритопы за счёт фенотипической и генетической изменчивости имеют широкую возможность перемещать оптимум вдоль градиентов факторов среды. Это положение во многом объясняется тем, что стенотопы чаще представлены видами «специалистами» с выраженным свойством S-стратегии (Grime, 1979, 1988; Миркин, 1985; Миркин и др., 1988) и потому их реализованная ниша ближе к фундаментальной. У эвритопов в большей мере выражены признаки R-стратегии и потому фундаментальная ниша всегда намного больше реализованной, что даёт им потенциальную возможность, при снижении уровня конкуренции, существовать в широких пределах градиентов факторов. Это положение является центральным тезисом фитоиндикации, позволяющим отбирать из большого числа видов наиболее тонкие индикаторы. Кроме традиционных индикационных задач, мониторинговый профиль позволяет рассматривать экологию опыления на уровне фитоценозов и, что очень важно, её трансформацию на хронотренде. Ещё в 1982 году К. Фегри и Л. Ван дер Пэйл (1982) среди направлений современной экологии указывали на важность изучения экологии опыления на уровне сообществ, что выводит нашу статью на высокий уровень актуальности.
МАТЕРИАЛЫ И МЕТОДЫ
Для проведения мониторинговых исследований в августе 1988 года на горе Опук был заложен модельный профиль (за 10 лет до официального создания заповедника). Специально планировался для ведения долгосрочного наблюдения за состоянием опукских экосистем (рис. 1). Ниже, мы последовательно (табл. 1), приводим характеристику площадок описания модельного профиля (рис. 2) от уровня моря (юго-восточное побережье) (точка 1) до северо-западного склона (точка 40). Повторное геоботаническое описание сообществ было выполнено в июле 2023 года. Подробно указываем место заложения профиля и локализацию площадок описаний в расчёте, что заинтересованные исследователи смогут продолжать наблюдения, и не только за состоянием растительных сообществ, а также и за другими компонентами фитоценозов и экосистем.
Первоначальное и последующее геоботаническое описание растительных сообществ модельного профиля выполнено согласно методическим рекомендациям (Голубев, Корженевский, 1985). Анализ способов и векторов опыления проведён по данным, извлечённым из базы данных «Экодата», созданной в Никитском ботаническом саду. Антэкологическая составляющая базы формировалась последовательно, причём в основу деления понятий была положена схема, предложенная Д. Франком и С. Клотцем (Frank, Klotz, 1990).
Все варианты опыления у растений опукского модельного профиля укладываются в четыре способа, каждый их которых имеет разное число векторов опыления. Так, в способе энтомогамия зафиксировано шесть векторов опыления: авто-анемо-энтомогамия, кейсто-авто-энтомогамия, апо-авто-энтомогамия, анемо-энтомогамия, клейсто-энтомогамия, авто-энтомогамия; в способе анемогамия – семь векторов: клейсто-энтомо-анемогамия, авто-энтомо-анемогамия, энтомо-авто-анемогамия, гидро-анемогамия, энтомо-анемогамия, клейсто-анемогамия, авто-анемогамия; в способе автогамия – три вектора: апо-автогамия, клейсто-автогамия, энтомо-автогамия; в способе гидрогамия известен лишь один вектор – авто-гидрогамия. В основу приведённой типизации положен структурно-функциональный подход по признакам агентов опыления – биотических и абиотических и частично по происхождению пыльцы.
Оценку условий экотопа, вернее климатопа, на площадках описаний выполняли по специально разработанной программе “Pover”, созданной на базе Excel, по ранее опубликованному алгоритму (Корженевский, 1990), содержащую унифицированную информацию о диапазоне размещения видов растений вдоль градиентов. Устанавливались минимальное и максимальное значения градаций, а также оптимумы для каждого средового фактора.
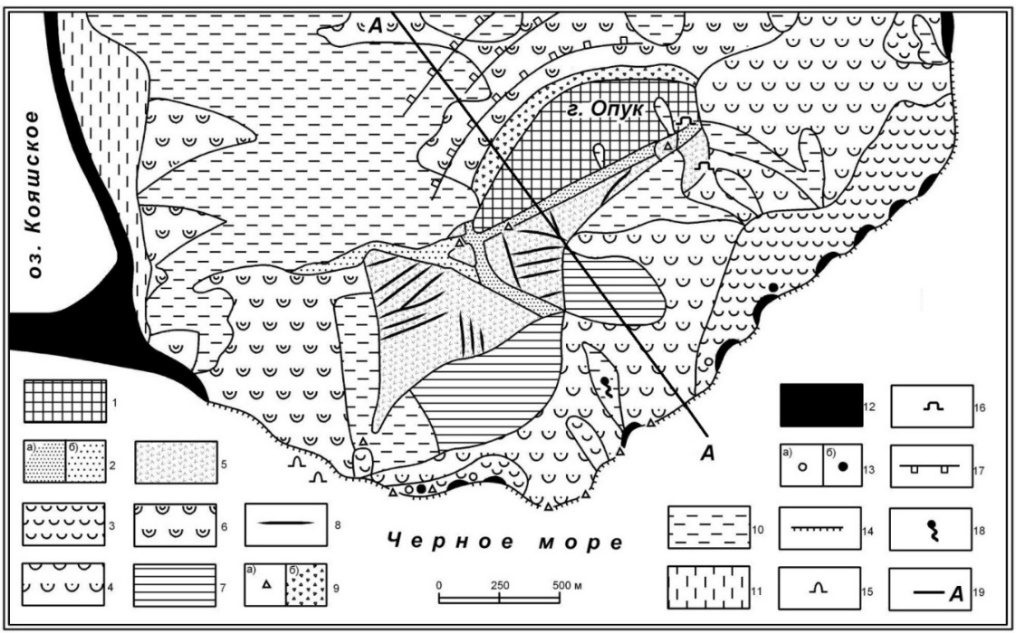
Рис. 1. Линия заложения модельного профиля и геоморфология горы Опук
1 – структурно-денудационное плиоценовое бронированное плато; 2 – сейсмогравитационные рвы (а – современный, б – позднечетвертичный); 3 – бугристо-холмистый и мелкоступенчатый рельеф современных оползней; 4 – грядово-ступенчатый рельеф современных-позднечетвертичных оползней; 5 – платообразно-ступенчатый рельеф современных оползней; 6 – холмисто-волнистый рельеф средне-позднечетвертичных оползней; 7 – депрессии оползневые и эрозионно-оползневые современные позднечетвертичные; 8 – оползневые и сейсмогравитационные трещины; 9 – обвалы и коллювиальные шлейфы (а – современные, б – позднечетвертичные); 10 – балки и ложбины современные и позднечетвертичные; 11 – делювиально-пролювиальные шлейфы современные и позднечетвертичные; 12 – пересыпи, пляжи и озёрные осушки современные; 13 – морская терраса нимфейская или новочерноморская (а – недеформированная, б – деформированная оползнями);
14 – клифы активные современные; 15 – гроты абразионные современные; 16 – штольни;
17 – террасы искусственные; 18 – источник; 19 –мониторинговый профиль.
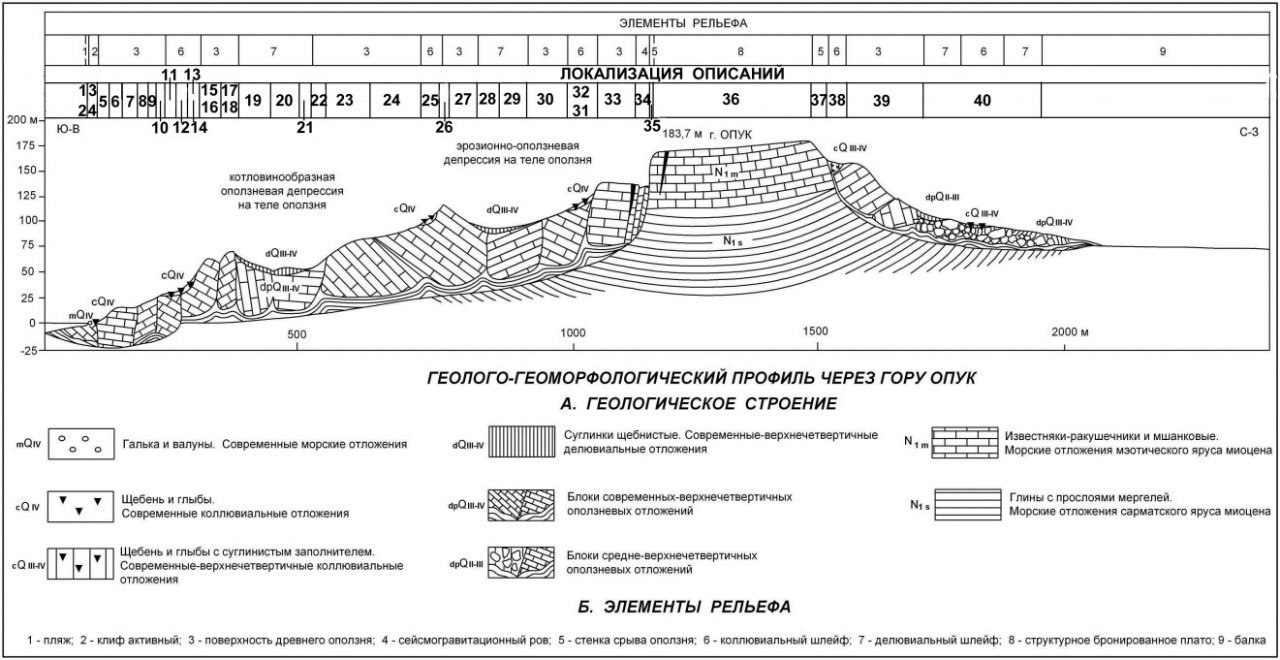
Рис. 2. Модельный профиль Опукского природного заповедника
Таблица 1
Топическая характеристика площадок геоботанических описаний мониторингового профиля
| № описания | Геоморфологический элемент рельефа | Высота над уровнем моря, м | Экспозиция, град. | Уклон,град. | Горная порода |
| 1 | 2 | 3 | 4 | 5 | 6 |
| 1 | Приморский склон (ПС), дикий пляж | 0–1 | 120 | 3 | Валуны и глыбы известняка |
| 2 | ПС, активный клиф высотой 5 м | 1–5 | 120 | 50–90 | Обвалившиеся глыбы известняка |
| 3 | ПС, мелкий современный коллювиальный шлейф | 5–7 | 100 | 25–30 | Глыбы и щебень известняка с песчаным заполнителем |
| 4 | ПС, старая обвальная ниша на отмирающем клифе, высотой до 10 м | 7–15 | 100 | 45 | Раздробленные слои и глыбы известняка оползневого блока |
| 5 | ПС, язык древнего стабильного оползня над клифом | 15–20 | 125 | 10–15 | Известняковый блок деляпсия с падением слоёв вглубь склона |
| 6 | ПС, язык древнего стабильного оползня (Q3-4) в 100 м от клифа | 15–20 | 125 | 10–15 | Известняковый блок деляпсия с падением слоёв вглубь склона |
| 7 | ПС, верховья оползневой депрессии | 20 | 60 | 4–5 | Мэотические известняковые слои с глыбами деляпсия |
| 8 | ПС, пологая ложбина | 20 | 60 | 4–5 | Известняковые слои с глыбами деляпсия |
| 9 | ПС, оползневой уступ | 20–25 | 165 | 15 | Известняк мэотический |
| 10 | ПС, эрозионно-оползневая депрессия, правый склон | 30–35 | 70 | 7–8 | Известняковые глыбы и блоки деляпсия стабильного древнего оползня |
| 11 | ПС, эрозионно-оползневая депрессия, днище | 30–35 | 130 | 6–7 | Известняковые глыбы и блоки деляпсия с коллювием |
| 12 | ПС, эрозионно-оползневая депрессия, левый склон с коллювиальный шлейфом | 30–35 | 180 | 20–25 | Известняковые слои с глыбами деляпсия с коллювием |
| 13 | ПС, тело древнего стабильного оползня, уступ оползневой террасы | 50–70 | 150 | 20 | Мшанковый сарматский известняк оползневого блока (dpQ3-4) |
| 14 | ПС, тело древнего стабильного оползня, поверхность оползневой террасы | 50–70 | 0 | 10 | Мшанковый сарматский известняк оползневого блока (dpQ3-4) |
| 15 | ПС, тело древнего стабильного оползня, эрозионно-оползневая депрессия глубиной 3-4 м | 70 | 0 | 18 | Мшанковый сарматский известняк оползневого блока (dpQ3-4) |
| 16 | ПС, тело древнего стабильного оползня, эрозионно-оползневая депрессия | 70 | 150 | 13 | Мшанковый сарматский известняк оползневого блока (dpQ3-4) |
| 17 | ПС, тело древнего стабильного оползня, оползневая псевдотерраса | 70–75 | 110 | 18 | Известняк оползневого блока (ИОБ) грубослоистый |
| 18 | ПС, тело древнего стабильного оползня, оползневой уступ | 70–75 | 0 | 60–70 | ИОБ грубослоистый |
| 19 | ПС, средняя часть, тело древнего стабильного оползня, эрозионно-оползневая депрессия глубиной 20-25 м, правый склон | 50–60 | 310 | 8 | ИОБ |
Таблица 1 (продолжение)
| 20 | ПС, средняя часть, тело древнего стабильного оползня, эрозионно-оползневая депрессия повышенная часть днища | 50–60 | 70 | 7 | ИОБ |
| 21 | ПС, средняя часть, тело древнего стабильного оползня, эрозионно-оползневая депрессия нижняя часть днища | 50–60 | 80 | 7 | ИОБ |
| 22 | ПС, средняя часть, тело древнего стабильного оползня, эрозионно-оползневая депрессия глубиной 20-25 м, левый склон | 50–60 | 180 | 15 | ИОБ |
| 23 | ПС, тело древнего стабильного оползня, уступ оползневой псевдотеррасы | 70–100 | 170 | 20–25 | ИОБ |
| 24 | ПС, тело древнего стабильного оползня, поверхность оползневой псевдотеррасы | 70–100 | 150 | 10 | ИОБ |
| 25 | ПС, тело древнего стабильного оползня, коллювиальный шлейф, уступ оползневой террасы с обвальными нишами | 100–115 | 190 | 18–20 | ИОБ |
| 26 | ПС, тело древнего стабильного оползня, коллювиальный шлейф, обвальная ниша | 100–115 | 190 | 70 | Известняк оползневого блока |
| 27 | ПС, верхняя часть, эрозионно-оползневая депрессия, структурный склон | 100–125 | 330 | 10 | ИОБ |
| 28 | ПС, верхняя часть, эрозионно-оползневая депрессия, правый склон | 100–125 | 330 | 10 | ИОБ, делювиальный суглинок |
| 29 | ПС, верхняя часть, эрозионно-оползневая депрессия, днище депрессии | 100–125 | 150 | 10 | ИОБ, делювиальный суглинок |
| 30 | ПС, верхняя часть, эрозионно-оползневая депрессия, левый склон с глыбами и смывом | 100–125 | 150 | 10 | ИОБ, делювиальный суглинок |
| 31 | ПС, верхняя часть, голова древнего оползня, коллювиальный шлейф | 115–130 | 150 | 22 | ИОБ с глыбами и щебнем |
| 32 | ПС, верхняя часть, голова древнего оползня, уступ оползневой террасы со стенкой обвальной ниши | 115–130 | 150 | 60–90 | ИОБ с глыбами и щебнем |
| 33 | ПС, верхняя часть, голова древнего оползня, верхняя оползневая терраса | 135 | 340 | 3–4 | Известняк мэотический (ИМ) оползневого блока |
| 34 | ПС, верхняя часть, голова древнего оползня, ров отседания под стенкой срыва | 125 | 50 | 5–20 | ИМ, глыбы коллювия поперечником до 3 м |
| 35 | ПС, верхняя часть, стенка срыва древнего оползня высотой до 40 м с тафони, нишами и полками | 137–175 | 140 | 80-90 | ИМ тонкослоистый, оолитовый, детритусовый и ракушечный |
| 36 | Структурное бронированной плато у вершины г. Опук с карровыми полями и трещинами бортового отпора | 175–180 | 155 | 1 | ИМ слоистый |
| 37 | Западный склон г. Опук, стенка срыва древнего оползня (Q2-3) высотой 10-15 м | 170–180 | 280 | 45–90 | ИМ слоистый |
| 38 | Западный склон г. Опук, голова древнего стабильного оползня, коллювиальный шлейф(Q3-4) | 150–160 | 300 | 15–20 | ИМ |
Таблица 1 (продолжение)
| 1 | 2 | 3 | 4 | 5 | 6 |
| 39 | Западный склон г. Опук, голова древнего стабильного оползня, оползневая терраса с уступом | 140–150 | 320 | 10–25 | Известняковые блоки и глыбы древнего деляпсия |
| 40 | Западный склон г. Опук, средняя часть, тело древнего стабильного оползня на поверхности, трансформированное античным производством | 120–140 | 310 | 5–10 | Известняковые блоки и глыбы древнего деляпсия |
Поскольку реакция видов в сообществе на градиенте фактора описывается колоколообразной кривой, приближающейся к теоретической кривой нормального распределения, то для неё приложимы методы расчёта эмпирического распределения, и в частности, асимметрии и эксцесса, позволяющие оценивать степень конкурентных отношений (эксцесс) и тенденции ресурсопотребления (асимметрия).
Оценка плотности основных опылителей цветковых растений проводилась методом учета насекомых с помощью кошения стандартным энтомологическим сачком с фиксированным количеством взмахов (Песенко, 1982).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Территориально Опукский природный заповедник размещается в Ленинском районе Республики Крым на землях бывших военного полигона и военсовхоза. В античную историческую эпоху на Опуке располагалось городище Киммерик, а прилегающая территория являлась его сельскохозяйственной округой – хорой (Клюкин, 2006). Природные особенности г. Опук давно привлекали внимание исследователей – геолога академика Н. И. Андрусова, ботаников Е. В. Вульфа, Е. В. Шифферс-Рафаловича, зоолога И. И. Пузанова и др., проводивших свои изыскания в первой половине ХХ века (Вахрушев, Вахрушев, 2006). С 1947 года гора Опук и острова Скалы-Корабли имели созологический статус – памятник природы, а с 1980 года – заповедное урочище. Опукский природный заповедник создан в 1998 году на площади 1592,3 га, в том числе 62 га акватории Чёрного моря.
Положение территории на юге умеренного пояса определяет значительное поступление солнечной энергии, а близость незамерзающего Чёрного моря оказывает смягчающее влияние на климат. Климат территории очень засушливый умеренно жаркий с очень мягкой короткой зимой и тёплым продолжительным летом. Средняя годовая температура воздуха уменьшается от Феодосии к Керченскому проливу от 11,7 до 10,6 °С. Самые холодные месяцы – январь и февраль, самые тёплые – июль и август. Средняя температура воздуха самого холодного месяца составляет около –0,5 °С, а самого тёплого +23,5 °С. Абсолютный минимум достигал –27 °С, а абсолютный максимум +38 °С. Безморозный период длится в среднем 217 дней.
Среднее количество атмосферных осадков составляет около 350 мм в год. Испаряемость превышает это значение примерно в 2,5 раза. В наиболее засушливые годы сумма атмосферных осадков снижается до 200 мм, а в наиболее влажные возрастает до 500–600 мм. Летом выпадают ливни, способные вызвать паводки высотой до 1,5 м в балках, оврагах и лощинах. Иногда слой суточных осадков превышает 100 мм. Засушливые периоды без дождей могут продолжаться более одного месяца. Устойчивый снежный покров отмечается только в суровые зимы. Высота снежного покрова достигает 35 см. Обычно снег ложится и стаивает несколько раз во время оттепелей. Постоянные водотоки отсутствуют. Ливневые и талые воды, изредка стекающие по эрозионным формам, задерживаются в прудах (аутах), которые в жаркое время года обычно пересыхают.
Поставив перед собой задачу, проследить изменения в структуре опылителей за прошедшие 35 лет на всех площадках геоботанических описаний, мы склонны считать, что видовое разнообразие, в том числе и среди опылителей, увеличивается в результате трансформации условий, а не их стабильности. Устойчивый состав сообществ, соответствующий «коридору толерантности» на градиентах факторов среды, оказывается, на самом деле, недостижимым, если местообитание подвержено частым нарушениям. Вызывать нарушения могут как однонаправленные современные экзогенные процессы, так и кратковременные катаклизмы.
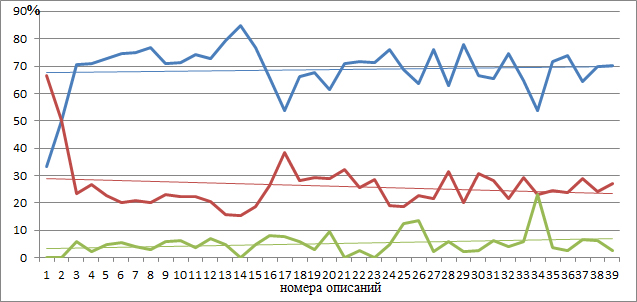
Конвертация числовых значений в проценты выполнена для сравнения относительного участия видов в процессах опыления на площадках описаний в 1988 и 2023 годах, отображения пропорций и выражения вероятностей. И хотя, в целом, кажущееся сходство процентного участия бросается в глаза на первый взгляд, то, на самом деле, все не так просто. Обратим внимание на мониторинговые площадки 9, 14, 15, 18, 28, 29, 30 и 37, здесь погодичная разница в структуре способов опыления более 10 %. При этом заметно снизилась роль энтомофильных растений и повысилась анемофильных. Как можно интерпретировать названные результаты? Во-первых, все эти перечисленные площадки в современном рельефе профиля находятся в условиях активно протекающих процессов рельефообразования, размещаясь на оползневых уступах, террасах, депрессиях и стенках срыва, а во-вторых, за прошедшие годы достаточно сильно сменился и видовой состав фитоценозов. Так в описании 9 в 1988 году было отмечено 68 видов растений, а стало 63, при этом убыло из состава сообщества 32 вида и внедрилось 27.
Несколько другая картина в описаниях 18 (первоначально было 26 видов, стало 56; исчезло 13, добавилось 43) и 37 (38/52, 18/32 соответственно). Конечно же, есть и такие описания, где в год повторного наблюдения отмечено увеличение численности энтомофильных видов растений и снижение анемофильных. Таким образом, можно констатировать, что наблюдаемые изменения это не что иное, как погодичная флуктуация, на фоне трансформации ландшафтно-экологических процессов в Крыму и глобальных климатических перестроек (Трансформация…, 2010). В какой-то степени это может быть и так, если не обратиться к графической демонстрации происходящих на мониторинговом профиле изменений процентных соотношений способов опыления в разные годы наблюдений (рис. 3).
На графиках (рис. 3) как в 1988, так и 2023 году, заметны отличия способов опыления на отдельных площадках геоботанических описаний и эти отличия обусловлены положением фитоценозов в рельефе. Выявить изменения позволяет линия тренда, проведённого через каждую кривую на графиках. В год закладки мониторингового профиля в приморской части (начало профиля) процент энтомофильных растений составлял 67,6 %, в то время как на северо-западном склоне (40 описание) 69,95 %; анемофильных – 29,2 и 23,3 %; автогамных – 3,3 и 6,8 %. Через 35 лет ситуация несколько поменялась: в начале профиля процент энтомофильных видов составил 72,8 %, а в конечном фитоценозе – 62,5 %; в тоже время анемофильных видов зафиксировано 22,8 и 30,8 %; автогамных – 4,47 и 6,7 %.
О чем говорят цифры и кривые графиков, да, изменился флористический состав на площадках описаний, и, естественно, изменились способы и векторы опыления. Конечно же, можно переложить всё на глобальное изменение климата, но мы пошли несколько дальше и методами фитоиндикации определили положение сообществ на градиентах факторов среды в год закладки профиля и год контроля.
Поскольку в эдафотопе за 35 лет особых изменений на градиентах факторов (режим увлажнения, рН субстрата, анионный состав и засоление, содержание карбонатов и азота, порозность субстрата) не произошло, мы не стали здесь приводить их графическую характеристику. В климатопе, попадающем под традиционное обсуждение происходящих и грядущих климатических изменений, особенно касаясь градиентов терморежим и криорежим, остановимся подробнее (рис. 4).
Наблюдения 1988 года
Наблюдения 2023 года
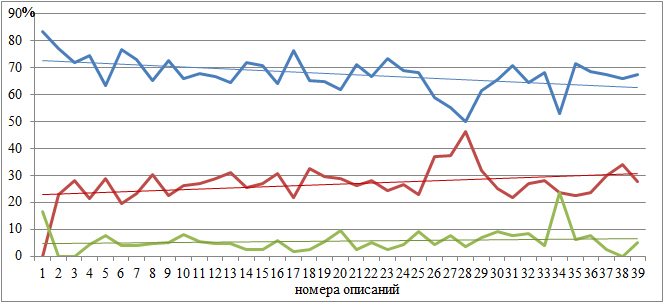
| энтомофильные виды | Линейная (энтомофильные виды) |
| анемофильные виды | Линейная (анемофильные виды) |
| автогамные виды | Линейная (автогамные виды) |
Рис. 3. Способы опыления растительных сообществ модельного профиля
Что реально показали фитоценозы в годы описания, если судить по линии тренда? В 1988 году среднеиюльская температура устойчиво повышалась от уровня моря на юго-восточном склоне до конечной точки на северо-западном (рис. 2).
В конкретных цифрах это выглядит так: начальная точка – +20,6 °С, финальная – +21,15 °С, отмечен рост температуры по мере удаления от береговой линии и пересечения массива горы Опук на 0,6 °С. Несколько другая картина наблюдалась в 2023 году: в начале профиля – +20,45 °С, в конце – +21,05 °С, то есть общий рост вдоль всего профиля 0,6 °С. Обобщим, за прошедшие 35 лет растительные сообщества индицируют незначительные изменения среднеиюльской температуры в сторону понижения: на 0,12 °С (начальная точка) и 0,10 °С (конечная).
Градиент терморежима, средняя температура июля
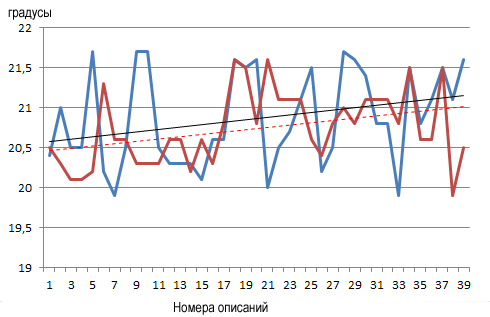
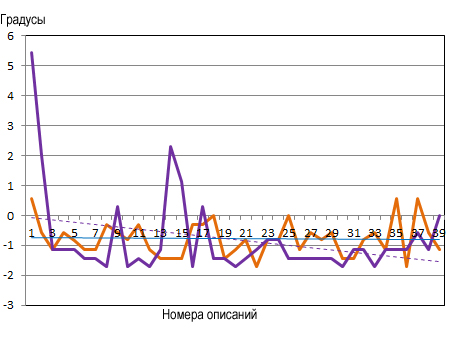
Градиент криорежима, средняя температура самого холодного месяца
| терморежим 1988 год | криорежим 1988 год |
| терморежим 2023 год | криорежим 2023 год |
| Линейная (терморежим 1988 год) | Линейная (криорежим 1988 год) |
| Линейная (терморежим 2023 год) | Линейная (криорежим 2023 год) |
Рис. 4. Положение сообществ на градиентах факторов среды.
Климатоп: терморежим и криорежим
Зимние температуры на хронотренде также несколько изменились (рис. 4). Прибрежная часть: –0,75 °С в 1988 году и –0,016 °С в 2023 году, общее потепление составило 0,73 °С; северо-западный склон: –0,80 °С (1988 г.) и –1,5173 °С (2023 г.), понижение на –0,75 °С. В 1988 году разница зимних температур между первой и последней точкой составляла –0,5 °С, а в 2023 году она снизилась до –1,5 °С, то есть на минус один градус за 35 лет. Что влияет на такие изменения климатических параметров ответить однозначно достаточно сложно и скорее всего, требует специального рассмотрения и обсуждения климатологов.
Анализ данных по учетам численности диких пчел – основных опылителей цветковых энтомофильных растений в различных природных зонах Крыма показал, что и на антропогенно измененных, и на заповедных территориях численность пчел за последние 30 лет существенно снизилась. В первом случае, в частности на территориях сельхозугодий, которые всегда отличались значительными флуктуациями численности насекомых, а также большом разнообразии состава посевов достичь достоверности оценок в отношении долговременных трендов изменения численности насекомых трудно. На участках же, где естественная растительность сохранилась относительно хорошо и, тем более, на заповедных территориях многолетние колебания численности насекомых имеют значительно меньшую амплитуду и результаты оценок численности насекомых в этом случае вызывают большее доверие. С равнение результатов учетов численности диких пчел на заповедных территориях Тарханкутского и Керченского полуостровов в
80-е и 90-е годы показали с таковыми в годы с 2000-го по 2024-й показали снижение плотности пчел с 9–16 до 2–5 экз./50 взмахов энтомологическим сачком. Более полному предоставлению этих данных будет посвящена специальная публикация. Причины столь резкого снижения численности пчел-опылителей не совсем понятны и требуют специальных исследований. Одно из предположений состоит в том, что снижение численности пчёл произошло из-за насыщения воздушной среды мельчайшими кусочками пластика, которые забивают трахеи пчел, приводя их к гибели. Это предположение согласуется с данным об общем снижении численности насекомых в природе в течение последних десятилетий в целом на Европейском континенте (https://en.wikipedia.org/wiki/Decline_in_insect_ populations).
ЗАКЛЮЧЕНИЕ
Временной промежуток в 35 лет, конечно же, для экосистемы заповедника Опук не так велик. К тому же, значительная удалённость от крупных населённых пунктов, вносит определённую лепту в сукцессионную динамику растительного покрова. Но, как бы то ни было, флористический состав поменялся, причём в ряде описаний более чем на половину, особенно в местах активного проявления современных экзогенных геоморфологических процессов. Условия на градиентах факторов среды эдафотопа практически не изменились, климатопа – с небольшими отклонениями, что требуют отдельного рассмотрения. А вот со структурой способов опыления ясно, что произошел десяти и более процентный сдвиг соотношения энтомогамии и анемогамии в сторону последней при более-менее стабильной автогамии. На наш взгляд, причиной этому стало снижение общей численности опылителей, вызванное антропогенными факторами, первостепенным из которых, по нашему предположению, является насыщение воздуха частицами микропластика, засоряющего дыхательные пути насекомых и приводящие их к гибели. Считаем необходимым продолжить мониторинговые наблюдения за опукскими экосистемами с повторами через 25 лет.
- Голубев В. Н., Корженевский В. В. Методические рекомендации по геоботаническому изучению и классификации растительности Крым. – Ялта, 1985. – 37 с.
- Вахрушев Б. А., Вахрушев И. Б. Заповедные объекты массива горы Опук и сопредельных территорий // Труды Никитского ботанического сада. – 2006. – Т. 126. – С. 23–33.
- Клюкин А. А. Природа и разнообразие факторов среды территории Опукского природного заповедника. Труды Никитского ботанического сада. – 2006. – Т. 126. – С. 8–22.
- Корженевский В. В. Об одном простом способе интерпретации экологических шкал // Экология. – 1990. – № 6.– С. 60–63.
- Иванов Е. С. Механизмы перехода энтомофильных культурных растений к самоопылению. Академия пчеловодства. Рыбное, 1995. – 15 с. Деп. в ВИНИТИ 25.10.1995, 2848-В95.
- Песенко Ю. А. Принципы и методы количественного анализа в фаунистических исследованиях. – М.: Наука, 1982. – 288 с.
- Плугатарь Ю.В., Корженевский В.В. Организация долгосрочного мониторинга на Главной гряде Крымских гор. I. Растительный покров // Научные записки природного заповедника «Мыс Мартьян». – 2017 – Вып. 8 – С. 26–60.
- Пономарев А. Н. Предмет и некоторые аспекты антэкологии // Вопросы антэкологии. – 1969. – С. 43–45.
- Фегри К., Ван дер Пейл Л. Основы экологии опыления. – М.: Мир, 1982. – 377с.
- Миркин Б. М. Теоретические основы современной фитоценологии. – М.: Наука, 1985. – 137 с.
- Миркин Б. М., Наумова Л. Г. Наука о растительности. – М.: Наука, 1998. – 564 с.
- Трансформация ландшафтно-экологических процессов в Крыму в ХХ веке – начале ХХI века. – Симферополь: Доля, 2010. – 304 с.
- Frank D., Klotz S. Biologisch-ökologische Daten zur Flora der DDR // Halle-Wittenberg: Wissenschaftliche Beitrage der Martin-Luther-Universität. – 1990. – P. 41. – 167 s.
- Grime J. P. Plant strategies and vegetation processes. – Chichester: Wiley and Sons, 1979. – 222 p.
- Grime J. P., Hodgson J. G., Hunt R. Comparative plant ecology: a functional approach to common British species. – London: Unwin Hyman, 1988. – 742 p.