
| Ekosistemy, 37: 65–72 (2024) | https://ekosystems.cfuv.ru |
| УДК 595.768.12-115.7 | DOI: 10.29039/2413-1733-2024-37-65-72 |
Многолетняя изменчивость рисунка переднеспинки колорадского жука Leptinotarsa decemlineata Say (Coleoptera, Chrysomelidae) на Среднем Урале
Галицкий А. В. 1, Джонсон Н. П. 1, Антипов А. В. 1, Гилев А. В. 1, 2
- Уральский федеральный университет имени Б. Н. Ельцина Екатеринбург, Россия
- Институт экологии растений и животных УрО РАН Екатеринбург, Россия
galickiiabc@mail.ru, brazajohnson@gmail.com, gilev@ipae.uran.ru
Популяции колорадского жука характеризуются высоким уровнем полиморфизма, что обеспечивает его высокую экологическую пластичность и способность к быстрой адаптации в различных условиях. В нашей работе рассматривается изменчивость рисунка центральной части переднеспинки колорадского жука Leptinotarsa decemlineata в выборках разных лет в двух локалитетах Среднего Урала. Описаны новые варианты рисунка, не отмеченные ранее на территории Европейской части Российской Федерации, для которых характерна более высокая степень меланизации. Для этих вариантов характерно слияние центральных полос между собой и с нижним пятном. В популяциях Среднего Урала доминировали варианты 1–3 с суммарной частотой встречаемости 32–55 %, в заметном количестве встречался новый вариант a (5–12 %). В сумме частота новых, более меланизированных вариантов составляла 17–36 %. Обнаружена хорошо выраженная многолетняя изменчивость частот вариантов рисунка переднеспинки, связанная с постепенным повышением частоты встречаемости массовых вариантов 1–3. Отмечено резкое повышение частоты варианта 9 в 1998 году, до 18–20 %, синхронно в обоих локалитетах. В целом синхронные выборки были более схожи друг с другом, чем с выборками из тех же локалитетов в другие годы. Синхронная временная динамика частот встречаемости вариантов окраски может свидетельствовать о принадлежности этих поселений к одной большой популяции колорадского жука. В целом полученные результаты свидетельствуют о продолжающихся процессах становления популяционной структуры вида в новом ареале расселения – на территории Среднего Урала.
Ключевые слова: колорадский жук, изменчивость окраски, многолетняя динамика, Средний Урал.
ВВЕДЕНИЕ
Колорадский картофельный жук Leptinotarsa decemlineata Say, несмотря на многолетние усилия ученых и практиков, специалистов по борьбе с вредителями, остается особо опасным вредителем картофеля и других пасленовых культур. Литература, посвященная колорадскому жуку, огромна, и число работ с каждым годом только растет.
Завезённый в начале XX века в Европу, найдя там практически неисчерпаемую кормовую базу, и не встретив естественных врагов, он с тех пор быстро и беспрепятственно расселяется в Евразии, занимая все новые территории (Кохманюк, 1983; Ясюкевич и др., 2007). Расселение вида в новом обширном ареале всегда сопровождается внутривидовой морфологической дифференциацией, становлением популяционной структуры, выработкой комплекса адаптаций к новым условиям обитания. Ярко выраженная изменчивость меланинового рисунка колорадского жука позволяет относительно легко получить большой сравнительный материал и провести достаточно глубокий анализ этих процессов. В связи с этим колорадский жук стал классическим модельным объектом для микроэволюционных исследований.
Одним из первых на изменчивость окраски жуков рода Leptinotarsa обратил внимание У. Тауэр (Tower, 1906). Расселение колорадского жука в инвазивном ареале вызвало новую волну интереса к изменчивости его окраски (Кохманюк и др., 1978; Кохманюк, 1982, 1983; Овчинникова и др., 1984; Фасулати, 1985; Климец, 1997; Зелеев, 2002; Паутова, Пак, 2008, 2011; Бабкина и др., 2017). Интенсивно изучалось адаптивное значение окрасочных морф в различных условиях, в том числе устойчивость к ядохимикатам (Климец, 1988; Беньковская и др., 2008; Удалов, Беньковская, 2010). Выявлено соответствие расположения пигментных пятен и мест прикрепления мышечных пучков (Присный, 1980). В целом ряде работ изучалась генетика окраски жука (Гриценко и др., 1998; Boiteau, 1994; Benkovskaya, Udalov, 2011).
На Урале колорадский жук появился в 70-х годах, с начала 80-х встречается повсеместно (Малоземов, 1989). Однако территория Урала и прилежащие территории изучены неравномерно (Беньковская и др., 2008; Паутова, Пак, 2008, 2011; Удалов, Беньковская, 2010; Удалов и др., 2010). Наша работа отчасти закрывает этот пробел и посвящена изучению фенотипической структуры и ее динамики в популяциях колорадского жука Среднего Урала.
Цель работы – изучить изменчивость меланинового рисунка переднеспинки в разновременных выборках колорадского жука из двух локалитетов Среднего Урала и выявить возможные направления многолетней динамики фенотипической структуры.
МАТЕРИАЛ И МЕТОДИКА
Выборки жуков были взяты в Свердловской области на частных садовых и придомовых участках в окрестностях села Первомайского (Нижнесергинский район) в 1997, 1998, 2000 годах, и в пригородах Екатеринбурга – поселок Кольцово в 1998 году, поселок Горный Щит в 2012 году (рис. 1). Выборки из поселков Кольцово и Горный Щит расположены близко друг к другу, можно считать их разновременными выборками из одной популяции. Расстояние между Екатеринбургом и селом Первомайским составляет около 65 км.
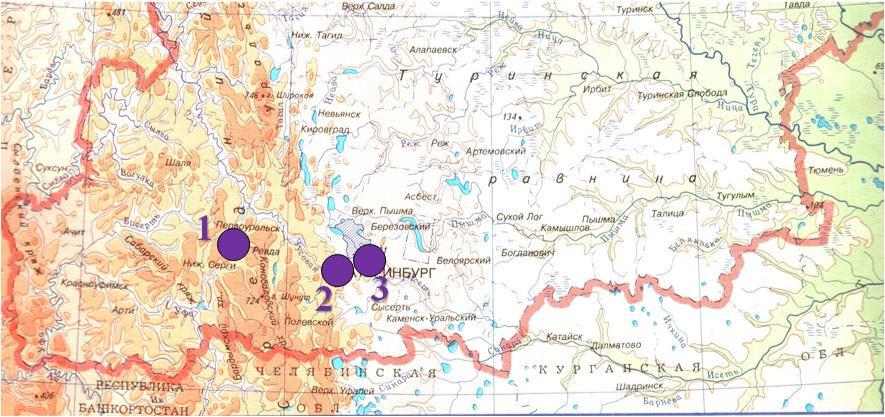
Рис. 1. Пункты сбора колорадского жука (южная часть Свердловской области)
1 – с. Первомайское; 2 – п. Горный Щит; 3 – п. Кольцово.
Все изученные выборки располагаются в пределах горно-таёжной полосы Среднего Урала, что существенно нивелирует возможное влияние ландшафтно-географических факторов. Объем собранного материала приведен в таблице 1.
Изменчивость рисунка переднеспинки жуков оценивали по схеме, предложенной С. Р. Фасулати (1985), с дополнениями. В этой схеме выделяется 9 вариантов рисунка (рис. 2, варианты 1–9) по степени развития центральной группы пятен (система пятен A, B и P по Кохманюк, 1982).
В изученных нами выборках были обнаружены варианты, не укладывающиеся в эту схему и отличающиеся более высокой степенью меланизации. Это проявлялось в слиянии центральных пятен в нижней части с образованием V-образной фигуры, а также в слиянии с ними нижней точки (рис. 2, варианты a–f). Отсутствие их у С. Р. Фасулати (1985) может
Таблица 1
Достоверность различий между выборками колорадского жука (критерий χ2)
| Кольцово, 1998 | Горный Щит,2012 | Первомайское,1997 | Первомайское,1998 | Первомайское,2000 | |
| Кольцово, 1998 (n=41) | — | 30,69** | 24,63* | 7,58 | 19,89 |
| Горный Щит, 2012 (n=173) | — | 35,81** | 95,29*** | 26,28* | |
| Первомайское, 1997 (n=143) | — | 42,81*** | 23,28 | ||
| Первомайское, 1998 (n=320) | — | 76,57*** | |||
| Первомайское, 2000 (n=400) | — |
Примечание к таблице. Звездочками показаны уровни достоверности различий: * – p<0,05; ** – p<0,01; *** – p<0,001.

Рис. 2. Основные типы рисунка центральной части переднеспинки имаго колорадского жука Leptinotarsa decemlineata Say (по Фасулати, 1985 с дополнениями)
1–9 – типы рисунка, выделенные Фасулати (1985); a–f – новые типы рисунка, обнаруженные в уральских популяциях.
означать их существенную редкость в европейской части ареала. На Среднем Урале частота этих вариантов часто превышает 40 %, поэтому мы сочли целесообразным выделить их и рассматривать отдельно. Обозначения наших вариантов даны таким образом, чтобы можно было легко добавить новые ещё более меланизированные. В целом схема изменчивости рисунка переднеспинки колорадского жука с нашими дополнениями теперь насчитывает 15 вариантов (рис. 2).
Достоверность различий между выборками по частотам встречаемости вариантов окраски оценивали с помощью критерия Хи-квадрат.
Для оценки уровня различий были рассчитаны фенотипические дистанции Кавалли-Сфорца, традиционно применяемые в таких исследованиях (Животовский, 1991). Матрицы дистанций обработаны методами многомерного неметрического шкалирования. Все расчеты выполнены в программах Microsoft Excel 2010 и Statistica v. 8.0 (StatSoft, Ink., 1984–2007).
РЕЗУЛЬТАТЫ
Спектры изменчивости в разновременных выборках колорадского жука из уральских популяций приведены на рисунке 3.
Видно, что все выборки сходны по своим спектрам изменчивости. Во всех выборках доминируют варианты 1–3 и 9. Это в целом совпадает с результатами С. Р. Фасулати (1985) и других авторов. В заметном количестве присутствует также более темный вариант а, для которого характерно слияние центральных пятен в нижней части (рис. 2, 3). В большинстве случаев, тем не менее, различия между выборками по критерию Хи-квадрат высоко достоверны (табл. 1).
Анализ сходства изученных выборок по частотам встречаемости вариантов окраски методами многомерного неметрического шкалирования показал, что наиболее близки между собой синхронные выборки из поселков Кольцово и Первомайское (1998 г.) (рис. 4).
По критерию Хи-квадрат эти выборки не различаются (табл. 1). Напротив, выборки разных лет из одного и того же пункта (п. Первомайское) хорошо отличаются (рис. 3б, 4). Хорошо различаются также и выборки 1998 и 2012 годов из окрестностей Екатеринбурга (рис. 3а, 4). Интересно отметить, что географические различия между удаленными пунктами (п. Первомайское и окрестностей Екатеринбурга) выражены заметно слабее (рис. 3, 4).
На рисунке 4 видно, что различия между разновременными выборками из п. Первомайское выражены преимущественно вдоль первой оси. Различия между выборками Кольцово (1998 г.) и Горный Щит (2012 г.) также выражены вдоль первой оси и, вероятно, отражают не географические, а хронографические различия (рис. 4). Таким образом, первую ось можно интерпретировать как отражающую резкие межгодовые колебания фенотипической структуры популяций. Любопытно, что вдоль второй оси выборки также располагаются упорядоченно по времени, от самой ранней, Первомайское (1997 г.), до самой поздней, Горный Щит (2012 г.) (рис. 4). Вторую ось, таким образом, тоже можно интерпретировать как отражающую изменения во времени, только более длительные и однонаправленные.
В целом данную картину можно интерпретировать, как наличие хорошо выраженной многолетней динамики фенотипических признаков в единой большой популяции. Эта динамика характеризуется постепенным повышением доли вариантов 1 и a (рис. 3). Резкий скачок 1998 года был связан с уменьшением доли варианта 1 и увеличением доли варианта 9.
К сожалению, мы не располагаем непрерывным рядом наблюдений и не можем сказать, является ли этот скачок чем-то исключительным, или же это достаточно обычное событие в динамике фенотипического состава популяции колорадского жука.
ОБСУЖДЕНИЕ
Расселение инвазивного вида в новом ареале – процесс, крайне интересный с точки зрения теории эволюции. Это чаще всего сопровождается фенотипической дифференциацией, происходит становление популяционной структуры в новой части ареала, формирование
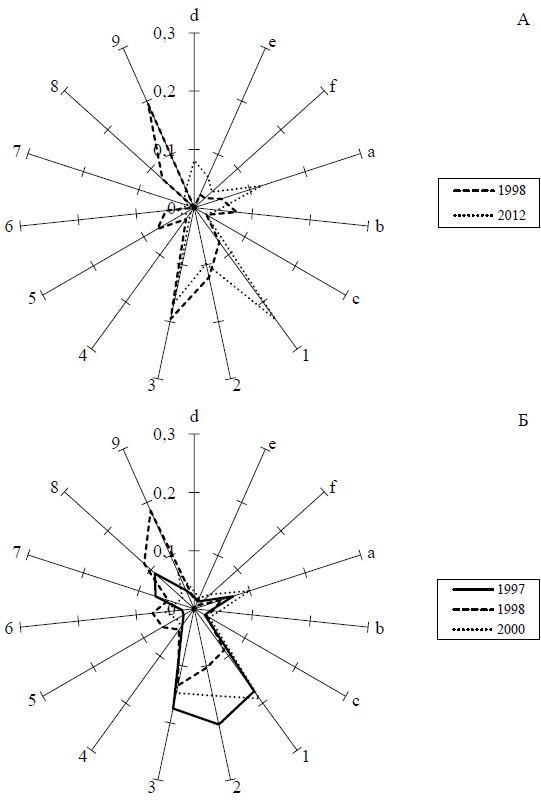
Рис. 3. Частоты встречаемости типов рисунка в выборках колорадского жука в популяциях Екатеринбурга (А) и Первомайского (Б)
нового комплекса адаптаций. Так, морфологическими и молекулярно-генетическими методами выявляется существенная внутривидовая дифференциация у колорадского жука, вплоть до подвидового уровня (Фасулати, 1985; Удалов, 2010; Фасулати, Иванова, 2022; Benkovskaya, Udalov, 2011; Insect Pests of Potato…, 2012). При этом популяции Старого Света генотипически мономорфны и представляют собой один подвид (Insect Pests of Potato…, 2012; Karagyan et al., 2021), но существенно дифференцированы фенотипически (Фасулати, 1985; Зелеев, 2002; Фасулати, Иванова, 2022; Benkovskaya, Udalov, 2011).
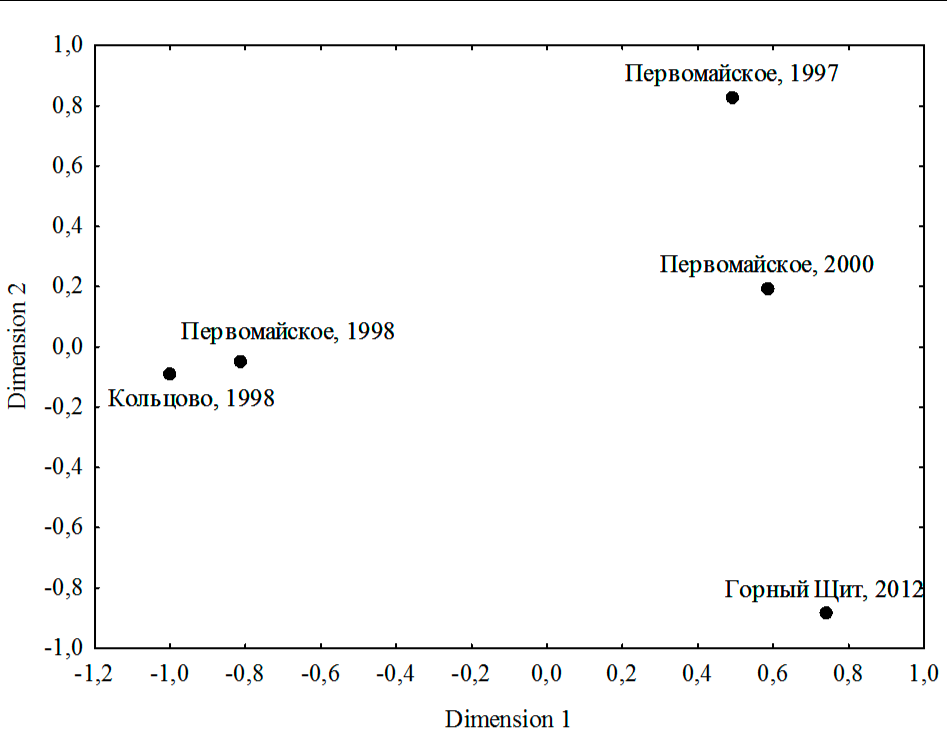
Рис. 4. Результаты анализа фенотипического сходства выборок колорадского жука методами многомерного неметрического шкалирования
По оценкам Ф. С. Кохманюка (1983), становление популяционной структуры у колорадского жука занимает период около 10 лет. За это время происходит формирование устойчивых локальных группировок, стабилизируется полиморфизм, снижается амплитуда колебаний частот морф (Кохманюк, 1983). Стабильность полиморфизма, высокая устойчивость частот морф в ряду лет характерна для многих популяций насекомых (Новоженов, Коробицын, 1972; Новоженов, 1980, 1989; Васильев, 1988, 2005). Как показали исследования А. Г. Васильева (2005), чаще всего размах внутрипопуляционной хронографической изменчивости существенно ниже уровня межпопуляционных различий.
Появление колорадского жука на Среднем Урале произошло в конце 70-х – начале 80-х годов (Малоземов, 1989). К настоящему времени, согласно Ф. С. Кохманюку (1983), формирование популяционной структуры жука уже должно в общих чертах завершиться, частота основных фенов – стабилизироваться. Резкие колебания частот все же возможны вследствие различного рода экстремальных воздействий. В целом наблюдаемая нами картина этим представлениям соответствует. Наблюдается медленная динамика фенотипической структуры на больших промежутках времени (в нашем случае – 15 лет), изредка происходят резкие отклонения частот фенотипов, такие как в 1998 году (рис. 3, 4). Фенотипическая структура популяции при этом существенно не меняется, доминируют одни и те же варианты рисунка.
Интересно отметить, что в Башкортостане в тот же период времени наблюдалась иная динамика частот морф, с 1994 по 2002 увеличилась частота вариантов 3, 6, 9, что авторы связывают с инсектицидными обработками и дифференцированным выживанием разных фенотипов (Удалов, Беньковская, 2010). Несомненно, инсектициды являются мощным фактором, способным существенно изменить структуру популяции. Неодинаковую устойчивость разных морф колорадского жука к пестицидам отмечали неоднократно (Климец, 1988; Удалов и др., 2010; Беньковская, Дубовский, 2020). Вместе с тем, это не единственный фактор, к которому колорадский жук должен адаптироваться, не менее важным и постоянно действующим фактором является температура, которая существенно ограничивает его распространение (Ясюкевич и др., 2007). Климат Среднего Урала заметно холоднее, что и может обуславливать различия фенотипической структуры популяций жука Южного и Среднего Урала.
Благодарности. Выражаем свою признательность С. В. Яковлевой, любезно предоставившей материал для анализа.
Список литературы
Бабкина Л. А., Балабина И. П., Тригуб Н. И., Миронов С. Ю., Мерзлякова К. В. Оценка фенотипического полиморфизма рисунка переднеспинки колорадского жука Leptinotarsa decemlineata Say в популяциях Курской области // Самарский научный вестник. – 2017. – Т. 6, № 1 (18). – С. 10–14.
Беньковская Г. В., Дубовский И. М. Распространение резистентности к инсектицидам в сибирских популяциях колорадского жука в связи с территориальной экспансией вредителя // Вестник защиты растений. – 2020. – 103 (1). – С. 37–39.
Беньковская Г. В., Удалов М. Б., Хуснутдинова Э. К. Генетическая основа и фенотипические проявления резистентности колорадского жука к фосфорорганическим инсектицидам // Генетика. – 2008. – Т. 44, № 5. – С. 638– 644.
Васильев А. Г. Эпигенетическая изменчивость: неметрические пороговые признаки, фены и их композиции // Фенетика природных популяций. – М., 1988. – С. 158–169.
Васильев А. Г. Эпигенетические основы фенетики: на пути к популяционной мерономии. – Екатеринбург: Академкнига, 2005. – 640 с.
Гриценко В. В., Глотов Н. В., Орлинский Д. Б. Эколого-генетический анализ изменчивости центральных элементов рисунка переднеспинки у колорадского жука (Leptinotarsa decemlineata) // Зоологический журнал. – 1998. – Т. 77, № 3. – С. 278–284.
Животовский Л. А. Популяционная биометрия. – М.: Наука, 1991. – 271 с.
Зелеев Р. М. Оценка полиморфизма рисунка переднеспинки и надкрылий колорадского жука, Leptinotarsa decemlineata, в окрестностях Казани // Зоологический журнал. – 2002. – Т. 81, № 3. – С. 316–322.
Климец Е. П. Выявление чувствительности колорадского жука к действию инсектицидов с помощью фенов // Фенетика природных популяций. – 1988. – С. 111–117.
Климец Е. П. Дискретные вариации рисунка на дорсальной стороне тела колорадского жука (Leptinotarsa decemlineata) // Популяционная фенетика. – 1997. – С. 45–58.
Кохманюк Ф. С. Изменчивость фенетической структуры популяций колорадского жука (Leptinotarsa decemlineata Say) в пределах ареала // Фенетика популяций. – 1982. – С. 233–243.
Кохманюк Ф. С. Становление структуры вида в новом ареале на примере колорадского жука // Экология. – 1983. – № 1. – С. 57–61.
Кохманюк Ф. С., Климец Е. П., Бибицкая Л. А., Пискунова В. В. Об изменчивости рисунка на переднеспинке колорадского жука // Физиологическая и популяционная экология животных. –1978. – Вып. 5 (7). – С. 141–148. Малоземов Ю. А. Динамика распространения колорадского жука на Урале // Всесоюзное совещание по
проблеме кадастра и учета животного мира: тез. докл. Ч. IV. – Уфа, 1989. – С. 181.
Новоженов Ю. И. Полиморфизм и непрерывная изменчивость в популяциях насекомых // Журнал общей биологии. – 1980. – Т. 41, № 5. – С. 668–679.
Новоженов Ю. И. Хронографическая изменчивость популяций // Журнал общей биологии. – 1989. – Т. 50, № 2. – С. 171–183.
Новоженов Ю. И., Коробицын Н. М. Аберративная изменчивость в природных популяциях насекомых // Журнал общей биологии. – 1972. – Т. 33, № 3. – С. 315–323.
Овчинникова Н. А., Климец Е. П., Маркелов Г. В. Динамика генетической структуры популяции колорадского жука на территории Липецкой области // Генетика. – 1984. – Т. 20, № 1. – С. 140–143.
Присный А. В. Морфологическая основа рисунка переднеспинки у колорадского жука (Leptinotarsa decemlineata) // Зоологический журнал. – 1980. – Т. 59, № 10. – С. 1575–1577.
Удалов М. Б. Формируются два подвида колорадского жука? // В мире научных открытий. – 2010. – № 4– 1 (10). – С. 74–76.
Удалов М. Б., Беньковская Г. В. Изменение уровня полиморфизма в популяциях колорадского жука на Южном Урале // Экологическая генетика. – 2010. – Т. 8, № 3. – С. 61–66.
Удалов М. Б., Беньковская Г. В., Хуснутдинова Э. К. Структура популяции колорадского жука на Южном Урале // Экология. – 2010. – № 2. – С. 126–133.
Фасулати С. Р. Полиморфизм и популяционная стратегия колорадского жука Европейской части СССР // Экология. – 1985. – № 6. – С. 50–56.
Фасулати С. Р., Иванова О. В. Изменчивость фенетической структуры и экологических адаптаций во вторичном ареале колорадского жука Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae) // Труды РЭО. – 2022. – Т. 93. – С. 52–68.
Ясюкевич В. В., Попова Е. Н., Гельвер Е. С., Ривкин Л. Е. Влияние климатических факторов на формирование ареала колорадского жука (Leptinotarsa decemlineata Say) // Проблемы экологического мониторинга и моделирования экосистем. – СПб.: Гидрометеоиздат, 2007.– С. 348–379.
Benkovskaya G. V., Udalov M. B. Colorado potato beetle investigations in the South Urals. – New York: Nova Science Publishers, Inc. 2011. – 54 p.
Boiteau G. Genetics of the Colorado potato beetle, Leptinotarsa decemlineata (Say) // Novel aspects of the biology of Chrysomelidae / [Jolivet P. H., Cox M. L., Petitpierre E. (eds)]. – Series Entomologica, vol 50. – Springer, Dordrecht, 1994. – P. 231–236.
Insect Pests of Potato: Global Perspectives on Biology and Management / [Edited by P. Giordanengo, Ch. Vincent and A. Alyokhin]. – Academic Press, 2012. – 614 p.
Karagyan G., Lukhtanov V., Mazmanyan M., Stepanyan I., Ghrejyan T., Abakumova A., Nesterova O. Genetic implications of a biological invasion: Chromosomal and DNA barcode monomorphism in Old World populations of Colorado potato beetle Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) // European Journal of Entomology. – 2021. – V. 118. – P. 344–354.
Tower W. L. An investigation of evolution in Chrysomelid beetles of the genus Leptinotarsa. – Wash.: Publ. Carnegie Inst., 1906. – 320 p.
Galitsky A.V., Dzhonson N.P., Antipov A.V., Gilev A.V. Long-term variability in the pronotum pattern of the Colorado potato beetle Leptinotarsa decemlineata Say (Coleoptera, Chrysomelidae) in the Middle Urals // Ekosistemy. 2024. Iss. 37. P. 65–72.
Populations of the Colorado potato beetle exhibit by a high level of polymorphism, which enables them to have high ecological plasticity and quickly adapt to various conditions. Our research examines the variability of the pattern of the central part of the pronotum of the Colorado potato beetle Leptinotarsa decemlineata in samples collected from two localities of the Middle Urals over several years. New variants of the pattern are described, which have not been previously observed in the European part of the Russian Federation. Higher degree of melanization is typical for these variations. Furthermore, these variants are characterized by the merging of the central stripes with each other and with the lower spot. In the populations of the Middle Urals, variants 1–3 dominated with a cumulative frequency of occurrence of 32–55 %; and a new variant a was found in significant quantities (5–12 %). In total, the frequency of newly registered variants with increased melanization was 17–36 %. A well-expressed long-term variability in the frequencies of variants of the pronotum pattern was discovered, associated with a gradual increase in the frequency of occurrence of mass variants 1–3. Synchronously in both localities, a sharp increase up to 18–20 % was recorded in the frequency of variant 9 in 1998. Overall, synchronous samples were more similar to each other than to samples from the same localities in other years. The synchronous temporal dynamics of variant color frequencies may prove that these settlements belong to one large population of the Colorado potato beetle. In general, the obtained results indicate the ongoing processes of population structure formation of the species in the new distribution area, specifically in the territory of the Middle Urals.
Key words: Colorado potato beetle, color variability, long-term dynamics, Middle Urals.
Поступила в редакцию 10.11.23
Принята к печати 16.01.24