
МИКРОЭЛЕМЕНТЫ В АГРОЦЕНОЗЕ ПЛОДОВОГО САДА ГРУШИ
ПРИ БИОЛОГИЗАЦИИ
TRACE ELEMENTS IN THE AGROCENOSIS OF A PEAR ORCHARD DURING BIOLOGIZATION
JOURNAL: «EKOSISTEMY», Issue 38, 2024
Publication text (PDF): Download
UDK: 634.13:574.4:579
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Klimenko O. E.1, Novitskaya A. P.1, Iakusheva N. N.2, Struchenko A. V.1, Fedyakhin I. A.1
Bondarenko Z. D., Bagrikova N. A.
1Federal State Budgetary Institute of Science «The Order of the Red Banner of Labour Nikita Botanical Gardens — National Scientific Center RAS», Yalta, Russia
2 Research Institute of Agriculture of Crimea, Simferopol, Russia
TYPE: Article
DOI: https://doi.org/10.29039/2413-1733-2024-38-35-48
PAGES: from 35 to 48
STATUS: Published
LANGUAGE: Russian
KEYWORDS: pear agrocenosis (Pyrus communis), trace elements, biologization, segetal vegetation, mixtures of sown grasses, microbial preparations, soil, plant nutrition.
.
ABSTRACT (ENGLISH):
The data of a two-factor field experiment conducted to research the content and mutual influence of trace elements (TEs) in soil, segetal vegetation, cultivated grasses and pear plants during the biologization of agrocenosis are presented. The experiment was carried out in the valley of the Salgir River in central Crimea (Simferopol region) on meadow alluvial carbonate soil in the orchard of pear (Pyrus communis L.) cv. Tavricheskaya on the rootstock VA 29. The experiment focused on the influence of the «sod» factor: 1) natural sod of the soil by segetal vegetation (SV) – control; 2) a mixture of herbs: Lolium multiflorum Lam. + Medicago sativa L. (HM2); 3) a mixture of herbs: L. multiflorum + M. sativa + Festuca pratensis Huds. + Trifolium pratense L. + Bromus inermis Leyss (HM4). Additionally, the effect of microbial preparations (MP) was investigated, which included: 1) a control group without MP, 2) Azotobacterin 07-Agro – the nitrogen-fixing and growth-stimulating agent; 3) Microbiocom-Agro (MBС) is a complex fertilizer with nitrogen-fixing, growth-promoting, phosphate-mobilizing, and biological protective properties. Mobile (available) forms of TEs: Fe, Mn, Cu, Zn and Co in soil, and total forms of the same elements in grasses and pear leaves were determined using the atomic absorption method. It was revealed that the concentrations of Fe, Mn and Co in the soil were below the regional background levels, while Mn and Co were at levels that were considered to below for fruit crops. The content of Cu and Zn in the soil was high, but did not exceed the maximum permissible concentration and was located in the following order: Mn ˃ Zn ˃ Fe ˃ Cu ˃ Co. Biologization caused an increase in the mobility of TE. MBC, when applied in combination with CT4, contributed to some relatively greater accumulation of Zn relative to Mn in the soil. It may cause Mn deficiency in the pear plant. The content of total forms of ME in herbs was generally optimal or high, but Mn was low. According to their content in herbs, TEs are arranged in the order: Fe ˃ Cu ≥ Zn ˃ Mn ˃ Co. The lack of Mn in grasses is associated with a deficiency of this element in the soil. MPs contributed to the accumulation of TEs in the biomass of sown grasses (except for Mn), which can be used for phytoremediation of soils contaminated with Co and Zn. The Mn content in herbs decreased under the influence of MBC. The concentration of total forms of TE (excluding Mn) in pear leaves was optimal and high, while Mn level was low. The series of element ratios was similar to that obtained for grasses, but Zn content in pear leaves was higher than Cu. The applied biologization methods increased the contents of Fe, Zn, and Co in leaves and decreased the concentration of Mn below the optimal level for pears. Herbs competed with the fruiting plants in the absorption of TEs, especially when their biodiversity increased and in combination with MPs. During the process of biologization the absorption of Co by the pear increased significantly and the absorption of Mn decreased, which caused a lack of the latter in the pear nutrition, both due to its low content in the soil and antagonism with Fe, Zn, Cu and Co in the plant. This manifested itself in external symptoms of deficiency, such as weak chlorosis and necrosis of young pear leaves. In general, the use of AB in conjunction with HM2 is the more optimal combination of sod and MP (from those studied) in terms of its effect on soil properties and nutrition of pear plants. A research should be conducted to determine optimal doses of manganese fertilizers for application through foliage for specific environmental conditions in order to improve the nutrition of pear by Mn at a low content of its mobile forms in carbonate soils and during biologization.
ВВЕДЕНИЕ
Микроэлементы (МЭ) играют важную роль в жизни растений. Они участвуют в процессах фотосинтеза, дыхания, окислительно-восстановительных реакциях, а также входят в состав ферментов. Многие МЭ обеспечивают устойчивость растений к неблагоприятным условиям среды. Обеспеченность сельскохозяйственных растений микроэлементами в том или ином агроценозе зависит от их содержания в почве в доступном для растений состоянии (Вернигорова и др., 2015), индивидуальных особенностей растения по поглощению и концентрации элемента в тканях, а также агротехники выращивания (Кабата-Пендиас, Пендиас, 1989; Карпухин, Бушуев, 2007; Кондаков и др., 2009). Однако многие МЭ являются и тяжелыми металлами и при определенных концентрациях в почве могут токсично воздействовать на сельскохозяйственные растения (Ильин, Сысо, 2001).
Применяемые приемы биологизации агроценоза плодового сада – введение дерново-перегнойной системы содержания почвы, а также биоудобрений на основе активных штаммов микроорганизмов могут значительно повлиять на содержание и доступность микроэлементов для плодовых культур. Нами ранее показано, что как задернение почвы различными травами, так и удобрение биопрепаратами, а также совместное их применение влияют на состояние, продуктивность и минеральное питание некоторых плодовых и декоративных растений (Клименко и др., 2019, 2020, 2024; Vystavna et al., 2020). Однако различные плодовые растения могут иметь разную потребность в МЭ и по-разному реагировать на введение того или иного приема биологизации (Трунов, Трунова, 2013; Sabir, 2013). Что касается агроценоза груши, то таких исследований нами в литературе не встречено.
Целью работы было изучить влияние приемов биологизации – задернения почвы различными злаково-бобовыми смесями трав и на их фоне применения биоудобрений – микробных препаратов различного спектра действия на содержание МЭ – Fe, Mn, Cu, Zn и Co в почве (подвижные формы), травах, используемых для задернения и листьях груши (валовые формы), их поглощение растениями и взаимное влияние.
МАТЕРИАЛ И МЕТОДЫ
Исследования проводили в 2023 году в ходе двухфакторного полевого опыта, заложенного в 2021 году в саду груши, расположенном в долине реки Салгир (с. Маленькое, Симферопольский р-он, Республика Крым). Сорт груши Таврическая на подвое айва ВА 29, схема посадки 3,5×1,5 м, формировка – стройное веретено. Закладка и проведение опыта осуществлялось согласно методике полевого опыта (Доспехов, 1985). Повторность опыта трехкратная. Размещение вариантов рендомизированное. Площадь элементарной делянки – 37 м2. Число деревьев на делянке – 5–7.
Факторами опыта были: задернение почвы сада многолетними травами и биоудобрения – микробные препараты (МП). Варианты опыта в факторе «задернение»:
1) естественное задернение (ЕЗ) сегетальной растительностью – контроль (рис. 1a);
2) задернение смесью трав: Lolium multiflorum Lam. и Medicago sativa L. (СТ2) в соотношении 1:1;
3) задернение смесью трав: L. multiflorum + M. sativa + Festuca pratensis Huds. + Trifolium pratense L. + Bromus inermis Leyss в соотношении 1:0,8:0,8:1:0,6 соответственно.
Травы высевали ручной сеялкой ListokLIE 09005. Норма высева семян 18–21 кг/га. Для создания дернового покрытия травы скашивали по мере отрастания на 30–40 см 3–4 раза за сезон и оставлением растительных остатков на месте в виде мульчи.
Фактор «МП» включал следующие варианты:
1) контроль – без МП;
2) биопрепарат Азотобактерин 07-Агро (АБ) – азотфиксатор, стимулятор роста;
3) Микробиоком-Агро (МБК) – комплексный препарат, обладающий азотфиксирующими, ростостимулирующими, фосфатмобилизующими и биопротекторными свойствами.
Препараты были предоставлены отделом сельскохозяйственной микробиологии НИИ сельского хозяйства Крыма (https://ckp-rf.ru/catalog/usu/507484/). МП вносили ежегодно весной после цветения груши.
Почва опытного участка – аллювиальная луговая карбонатная тяжелосуглинистая на современном глинистом аллювии. Перед закладкой опыта почва имела в метровом слое

Рис. 1. Задернение сада груши (вариант – естественное задернение) (a) и признаки недостатка Mn у груши сорта Таврическая – слабый хлороз и некроз молодых листьев (б)
(Крым, Симферопольский район, 2023 г.) (фото О. Е. Клименко)
следующие показатели: рНн2о 7,1–8,0, общих карбонатов – 7–12 %, гумуса 3,3–4,4 %, нитратного азота – 20–28 мг/кг, подвижного фосфора – 25–45 мг/кг и обменного калия – 310–406 мг/кг (Плугатарь и др., 2019). В целом почва была достаточно обеспечена элементами питания.
Образцы надземной биомассы трав отбирали перед кошением в апреле в трехкратной повторности методом пробных площадок (Казеев и др., 2016). Листья груши для анализа отбирали со средней части однолетних побегов из середины кроны по периметру дерева в фазу окончания интенсивного роста побегов (начало августа) в трехкратной повторности по 100 листьев. Подготовку образцов к анализу проводили согласно (Методические указания…, 1993).
Отбор образцов почвы для анализа проводили ежегодно в конце июля по 20-ти сантиметровым слоям до глубины 60 см в области ризосферы растений в трехкратной повторности. МЭ: Fe, Mn, Cu, Zn и Co экстрагировали из почвы ацетатно-аммонийным буфером с рН 4,8 при соотношении почва–раствор 1:10 согласно ДСТУ 4770:2007. Для подготовки к определению валовых форм МЭ образцы трав и листьев груши озоляли смесью Н2SO4 и H2O2 (Cоловьев, 1973). Определение содержания МЭ в вытяжках из почв и подготовленных растительных образцах проводили атомно-абсорбционным методом на анализаторе Квант-2МТ.
Статистическую обработку результатов выполняли методами дисперсионного и корреляционного анализов (Доспехов, 1985), используя пакет программ Statistica 07. Приведены средние значения (М), значения наименьшей существенной разницы (НСР) и коэффициенты парной корреляции (r) при 95 % доверительном уровне (t≤0,05).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Содержание подвижных форм МЭ, которые непосредственно доступны растениям, в 60-ти см слое почвы под садом было невысоким по Fe, Mn и Co, что было ниже регионального фона и по последним двум элементам соответствовало низкой обеспеченности почвы для культур повышенного выноса МЭ, к которым относятся и плодовые растения (Методические указания…, 1993; Скляр и др., 2021). Это во многом связано с карбонатностью почв и щелочной реакцией почвенного раствора (рис. 2, табл. 1).
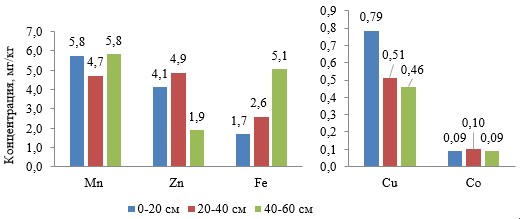
Рис. 2. Распределение подвижных форм микроэлементов в гумусовом горизонте луговой аллювиальной почвы (2023 год, цветом выделены слои разного уровня залегания)
Содержание Cu и Zn в почве было повышенным относительно фонового, особенно в верхнем слое почвы (рис. 2, табл. 1), что соответствовало средней и высокой обеспеченности почвы этими элементами, однако было ниже ПДК (табл. 1).
Таблица 1
Фоновое содержание и уровень обеспеченности почв Крыма МЭ (подвижные формы)
(по Ильин, Сысо, 2001 и Скляр и др., 2021)
|
Элемент |
Фоновое содержание |
Уровень обеспеченности для культур повышенного выноса МЭ |
ПДК |
||
|
низкий |
средний |
высокий |
|||
|
Mn |
23 |
<10 |
10–20 |
˃20 |
140 |
|
Zn |
0,45 |
<2 |
2–5 |
˃5 |
23 |
|
Cu |
0,39 |
<0,2 |
0,2–0,5 |
˃0,5 |
3 |
|
Co |
0,22 |
<0,15 |
0,15–0,30 |
˃0,30 |
5 |
Повышенное содержание Cu и Zn в почве может быть обусловлено многолетним использованием почв долины реки Салгир под сады и поступлением этих МЭ с пестицидами и минеральными удобрениями (Иванова, 2002; Лисецкий, Зеленская, 2023). Согласно уровню загрязнения почвы ТМ содержание этих элементов в почве контроля находилось на уровне фонового, установленного в результате длительного мониторинга содержания МЭ в почвах Крыма (Скляр и др., 2021).
В верхнем 60-ти см слое почвы МЭ распределяются не одинаково (рис. 2). Так, содержание Mn и Co не имело значительных колебаний в пределах этого слоя, тогда как Zn незначительно накапливался в слое 20–40 см. На глубине 40–60 см его содержание резко снижалось (в 2,6 раза) по сравнению с поверхностным слоем. Концентрация подвижных форм Cu была максимальной в слое 0–20 см и уменьшалась в слое 20–60 см в 1,5–1,7 раза, что связано с совмещенным механическим, сорбционным, карбонатным и геохимическим барьером, каковым является почва (Горбов, Безуглова, 2020). Содержание подвижного Fe, наоборот, постепенно возрастало с глубиной, что определяется высоким его содержанием в почвообразующей породе и значительным потреблением этого элемента растительностью из верхних слоев почвы.
Применяемые приемы биологизации агроценоза груши влияли на содержание подвижных форм МЭ в почве. Исследования показывают, что содержание подвижного Fe в верхнем 60-ти см слое почвы было низким, как в контроле, так и по вариантам опыта (табл. 2) и было ниже регионального фона – 7–15 мг/кг (Акчурин, Костенко, 2008).
Таблица 2
Влияние биологизации на содержание микроэлементов в почве (подвижные формы (мг/кг), среднее в слое 0–60 см, сад груши, 2023 г.)
|
Вариант |
Контроль |
АБ |
МБК |
Среднее по фактору «задернение» |
|
Fe |
||||
|
ЕЗ |
3,11 |
2,31 |
2,09 |
2,51 |
|
СТ2 |
2,09 |
2,88 |
3,49 |
2,82 |
|
СТ4 |
2,08 |
4,40 |
5,33 |
3,94* |
|
Среднее по фактору «МП»; НСР05 = 0,50; НСР05ч.с. = 0,86 |
2,43 |
3,20* |
3,64* |
|
|
Mn |
||||
|
ЕЗ |
5,44 |
4,91 |
5,27 |
5,21 |
|
СТ2 |
5,80 |
5,51 |
6,65 |
5,99* |
|
СТ4 |
7,20 |
6,78 |
6,48 |
6,82* |
|
Среднее по фактору «МП»; НСР05 = 0,61; НСР05ч.с. = 1,05 |
6,15 |
5,73 |
6,13 |
|
|
Zn |
||||
|
ЕЗ |
3,64 |
5,04 |
4,03 |
4,23 |
|
СТ2 |
6,23 |
8,70 |
8,43 |
7,78* |
|
СТ4 |
8,08 |
11,01 |
11,64 |
10,24* |
|
Среднее по фактору «МП»; НСР05 = 2,25; НСР05ч.с. = 3,90 |
5,98 |
8,25* |
8,03 |
|
|
Cu |
||||
|
ЕЗ |
0,59 |
0,73 |
0,56 |
0,62 |
|
СТ2 |
0,77 |
1,17 |
0,99 |
0,97* |
|
СТ4 |
0,78 |
1,37 |
1,38 |
1,18* |
|
Среднее по фактору «МП»; НСР05 = 0,35; НСР05ч.с. = 0,61 |
0,71 |
1,09* |
0,98 |
|
|
Co |
||||
|
ЕЗ |
0,092 |
0,090 |
0,096 |
0,093 |
|
СТ2 |
0,103 |
0,110 |
0,120 |
0,111* |
|
СТ4 |
0,126 |
0,131 |
0,125 |
0,127* |
|
Среднее по фактору «МП»; НСР05 = 0,012; НСР05ч.с. = 0,021 |
0,107 |
0,110 |
0,114 |
|
Примечание к таблице. Здесь и далее в таблицах НСР05 – для фактора «задернение» и «МП», НСР05 ч.с. – для частных средних; * – различия с контролем и ЕЗ по соответствующему фактору значимы (р<0,05). АБ – азатбактерии; МБК – Микробиоком-Агро.
Влияние задернения сеяными травами и МП по отдельности снижало его концентрацию в почве по сравнению с контролем по ЕЗ, а вот совместное влияние СТ4 с АБ и МБК приводило к существенному увеличению его содержания на 1,29 и 2,22 мг/кг или 41 и 71 % соответственно. Что касается влияния факторов, то задернение СТ4 достоверно увеличивало содержание Fe в 60-ти см слое почвы, доля влияния фактора составила 28 % общей дисперсии. По фактору МП оба препарата увеличивали содержание железа в почве, в большей мере МБК, доля влияния фактора составила 19 %, совместное влияние факторов оказалось наибольшим – 41 % общей дисперсии. Однако содержание подвижного Fe в почве оставалось низким.
Концентрация подвижного Mn в гумусовом горизонте луговой аллювиальной карбонатной почвы также была на уровне фоновой и находилась в пределах низкой обеспеченности для плодовых культур (табл. 1). Из применяемых приемов только задернение как фактор увеличивало содержание подвижного Mn на 0,78–1,61 мг/кг, максимально СТ4 (табл. 2). Доля влияния фактора составила 54 % в общей дисперсии. Внесение в почву микробных препаратов на фоне ЕЗ создавало тенденцию к снижению содержания Mn в почве. Это связано с окисляющей ролью бактерий, а также потреблением Mn самими микроорганизмами (Стойлов, 1967).
Применение МП по фону смеси трав, наоборот, увеличивало содержание подвижного Mn в почве, существенно на вариантах СТ2 с МБК и СТ4 с АБ на 1,21–1,34 мг/кг (22–25 %) от контроля по ЕЗ. Вероятно, корневые экссудаты сеяных трав не только удовлетворяют потребность бактерий в Mn, но и способствуют переведу валового Mn почвы в подвижное состояние.
Концентрация подвижных форм Zn в гумусовом слое контрольного варианта по ЕЗ была на порядок выше фоновой и находилась на среднем уровне обеспеченности плодовых культур (табл. 1, 2). Применяемые приемы как факторы способствовали увеличению подвижности Zn, особенно фактор «задернение», при воздействии которого содержание Zn в почве увеличивалось в 1,8–2,4 раза, максимально под действием СТ4 (табл. 2). Доля влияния фактора в общей дисперсии составила 56 %. МП как фактор в меньшей мере влияли на этот показатель и только АБ достоверно увеличивал содержание Zn на 2,27 мг/кг или на 38 % от контроля. Что касается совместного влияния приемов, то все они существенно повышали его концентрацию в почве на 5−8 мг/кг, максимально – сочетание СТ4 м МБК, при котором содержание подвижного Zn в почве оказывалось на уровне умеренного загрязнения – 10,1–15,0 мг/кг (Ильин, Сысо, 2001) в среднем для слоя 0–60 см, а в слое 20–40 см достигало 15–16 мг/кг, что в 35 раз превышало региональный фоновый уровень, хотя и было намного ниже ПДК (табл. 1). Это, вероятно, связано с антропогенным накоплением Zn с фунгицидами (La Pera et al., 2008) и с переводом его в подвижное состояние в результате действия трав и МП.
Содержание подвижной Cu в почве было на уровне фона и на высоком уровне обеспеченности растений этим элементом в контроле по ЕЗ. Оба изучаемых фактора и их сочетания способствовали увеличению подвижности Cu в гумусовом горизонте. Доля влияния фактора «задернение» составила 30 %, «МП» – 14 % общей дисперсии. Из частных средних оба МП на фоне СТ4 максимально и в равной мере увеличивали содержание подвижной Cu в почве в 2,3 раза по сравнению с контролем по ЕЗ (табл. 1).
Концентрация подвижного Co в почве была низкой и на порядок ниже регионального фона. Применяемое «задернение» как фактор достоверно увеличивало его содержание, хотя оно оставалось на низком уровне обеспеченности груши этим элементом. Из факторов только «задернение» способствовало существенному увеличению содержания доступного Со в почве, в большей мере СТ4. Доля влияния этого фактора составила 63 % общей дисперсии. Из частных средних наибольшему увеличению концентрации Cо в почве способствовал АБ на фоне СТ4 (табл. 2).
В целом содержание подвижных форм МЭ в почве контроля расположилось в следующий убывающий ряд: Mn ˃ Zn ˃ Fe ˃ Cu ˃ Co. Применяемые приемы биологизации, особенно применение МБК на фоне СТ4 способствовало некоторому относительно большему накоплению Zn по отношению к Mn.
Таким образом, применяемые приемы влияли положительно на подвижность и доступность растениям груши основных МЭ, которые в щелочных почвах малодоступны растению. При этом большее влияние на увеличение содержания МЭ в почвах оказывало «задернение» как фактор, который обеспечивал от 30 до 60 % общей дисперсии. Из трав СТ4 отличался большим воздействием на содержание всех МЭ в почве по сравнению с СТ2, что может свидетельствовать о положительном влиянии увеличения биоразнообразия трав при задернении. МП показывали некоторую специфичность в изменении содержания МЭ в почве. Так, на содержание Fe и Zn большее положительное влияние оказывал МБК в сочетании с СТ4, на Mn и Co – АБ, на содержание Сu в почве оба препарата влияли одинаково положительно на том же фоне трав. Некоторые опасения вызывает значительное увеличение подвижности цинка в почве под действием биологизации, что может вызвать увеличение его в тканях плодового растения и нарушения в минеральном питании МЭ.
Содержание МЭ в травах, участвующих в задернении почвы во многом зависит от количества доступных форм элемента в почве, биологических особенностей вида, а также агротехники. Данные показывают, что содержание валового Fe в травах, как в контроле по ЕЗ, так и по вариантам «задернения» было высоким, что превышало содержание подвижных его соединений в почве в 40–60 раз (табл. 2 и 3) и находилось в пределах оптимального для большинства луговых трав (Церлинг, 1990). Это свидетельствует о доступности других форм Fe в почве для травянистой растительности. В биомассе сеяных злаково-бобовых трав оно было достоверно выше в среднем по фактору на 25–27 мг/кг по сравнению с ЕЗ за счет высокого содержания Fe в бобовых растениях. МП как фактор также положительно влияли на накопление Fe в травах, максимально АБ. Доля влияния фактора «МП» была более значительной (32 %), чем задернения (21 %). Доля совместного влияния факторов составила 47 %, что свидетельствует о синергическом эффекте задернения и МП.
Таблица 3
Влияние биологизации на валовое содержание микроэлементов (мг/кг) в травах, задерняющих почву в агроценозе груши (2023 г.)
|
Вариант |
Контроль |
АБ |
МБК |
Среднее по фактору «задернение» |
|
Fe |
||||
|
ЕЗ |
137 |
201 |
130 |
156 |
|
СТ2 |
152 |
188 |
204 |
181* |
|
СТ4 |
169 |
176 |
203 |
183* |
|
Среднее по фактору «МП»; НСР05 = 1;НСР05ч.с. = 2 |
152 |
188* |
179* |
|
|
Mn |
||||
|
ЕЗ |
5,02 |
4,68 |
4,53 |
4,74 |
|
СТ2 |
5,52 |
7,58 |
5,40 |
6,17* |
|
СТ4 |
5,30 |
5,68 |
4,98 |
5,32* |
|
Среднее по фактору «МП»; НСР05 = 0,19; НСР05ч.с. = 0,33 |
5,28 |
5,98* |
4,97 |
|
|
Zn |
||||
|
ЕЗ |
34,4 |
44,0 |
50,8 |
43,1 |
|
СТ2 |
73,6 |
60,0 |
50,1 |
61,2* |
|
СТ4 |
31,7 |
48,7 |
42,7 |
41,1* |
|
Среднее по фактору «МП»; НСР05 = 0,01; НСР05ч.с. = 0,02 |
46,6 |
50,9* |
47,9* |
|
|
Cu |
||||
|
ЕЗ |
17,2 |
11,3 |
11,5 |
13,3 |
|
СТ2 |
41,8 |
31,0 |
40,0 |
37,6* |
|
СТ4 |
28,3 |
67,6 |
52,1 |
49,3* |
|
Среднее по фактору «МП»; НСР05 = 0,22; НСР05ч.с. = 0,38 |
29,1 |
36,6* |
34,5* |
|
|
Co |
||||
|
ЕЗ |
0,87 |
1,16 |
0,86 |
0,96 |
|
СТ2 |
1,24 |
3,63 |
2,57 |
2,48* |
|
СТ4 |
1,79 |
2,54 |
2,47 |
2,27* |
|
Среднее по фактору «МП»; НСР05 = 0,02; НСР05ч.с. = 0,03 |
1,30 |
2,44* |
1,97* |
|
Пояснение к таблице. АБ – азотобактерии; МБК – Микробиоком-Агро.
Корреляционный анализ данных показал, что содержание Fe в травах в условиях биологизации достоверно связано с концентрацией его в почве (r=0,46, р<0,05), а также с содержанием подвижных форм Zn и Cu в почве (табл. 4). Это может способствовать значительному улучшению питания растений железом при низкой его концентрации в почве
Таблица 4
Коэффициенты корреляции между содержанием МЭ в почве (подвижные формы) и их концентрацией в травах и листьях груши (n=27)
|
Элемент (почва) |
Травы |
Листья груши |
||||||||
|
Zn |
Cu |
Fe |
Mn |
Co |
Zn |
Cu |
Fe |
Mn |
Co |
|
|
Zn |
0,02 |
0,72 |
0,55 |
0,31 |
0,66 |
0,27 |
−0,16 |
0,40 |
−0,26 |
0,77 |
|
Cu |
0,08 |
0,62 |
0,49 |
0,30 |
0,60 |
0,28 |
−0,23 |
0,25 |
−0,30 |
0,82 |
|
Fe |
−0,17 |
0,67 |
0,46 |
0,04 |
0,47 |
−0,13 |
−0,46 |
−0,03 |
−0,59 |
0,86 |
|
Mn |
−0,22 |
0,57 |
0,24 |
0,04 |
0,35 |
0,07 |
0,10 |
0,56 |
0,02 |
0,49 |
|
Co |
−0,10 |
0,68 |
0,36 |
0,22 |
0,55 |
0,24 |
0,04 |
0,57 |
−0,09 |
0,48 |
Примечание к таблице. Коэффициенты корреляции более /0,46/ значимы (р<0,05).
и увеличению биомассы трав в агроценозе плодового сада (Клименко и др., 2024). Однако, высокое содержание Fe (150–250 мг/кг), в то же время, может вызвать снижение потребления Mn и нарушение соотношения Fe/Mn (Alvarez-Tinaut et al., 1980).
Содержание Mn в травах было низким, близким к содержанию его подвижных форм в почве, и связано положительно с изменениями его концентрации в почве при биологизации – достоверно увеличивалось под действием фактора задернение и под влиянием АБ (табл. 1 и 2). Доля влияния фактора «задернение» на содержание Mn в травах составила 23 %, МП – 14 %, совместное влияние факторов было значительным – 54 % общей дисперсии. Тенденция к снижению содержания Mn в травах под действием МБК может быть связана с дополнительным поглощением этого элемента микроорганизмами, входящими в комплекс. При низком содержании Mn в почве это может вызвать его недостаток в травах, а также в растениях груши.
Концентрация Zn и Cu в скошенной биомассе трав в контроле в 10–11 раз превосходила таковую в почве, что свидетельствует о значительной потребности растений в этих элементах для биосинтеза (табл. 2, 3), и была на уровне оптимальной (Zn) и высокой (Сu) для трав (Церлинг, 1990). Содержание Zn в травах под действием задернения СТ2 увеличивалось в 1,4 раза относительно ЕЗ, по фону СТ4, наоборот, существенно снизилось на 2 мг/кг. Доля влияния фактора составила 57 % общей дисперсии. Оба МП способствовали достоверному увеличению концентрации Zn в травах, в большей мере АБ. Из частных средних Zn максимально накапливался в травах при сочетании СТ2 с АБ в 1,7 раза относительно контроля, что может иметь важное значение при фиторемедиации антропогеннозагрязненных почв этим элементом. В целом, содержание Zn в травах не зависело от его количества в почве (табл. 4).
Задернение и МП как факторы способствовали накоплению Cu в травах. Доля влияния «задернения» была максимальной и составила 70 % общей дисперсии. Наибольшее накопление этого элемента отмечено на смеси СТ4, что в 4 раза превышало его содержание в травах по ЕЗ. Это также может быть использовано при ремедиации почв при повышенном содержании Сu в почве. Установлена положительная достоверная корреляционная зависимость между содержанием Cu в травах и концентрацией ее подвижных форм в почве (r=0,62), кроме того, она положительно связана с содержанием всех исследованных МЭ в почве при биологизации (табл. 4).
Установлено высокое содержание Со в травах в контроле по ЕЗ, несмотря на низкое содержание подвижного Со в почве (табл. 2 и 3). Применяемые приемы биологизации и их сочетания способствовали увеличению содержания Со в травах, причем доля влияния фактора «задернение» была более значительной (55 %) по сравнению МП (27 %). В большей степени положительно на содержание этого элемента повлияло сочетание СТ2 и АБ – увеличивало в 4,2 раза относительно контроля по ЕЗ. Содержание Со в травах положительно достоверно коррелировало с концентрацией его подвижных форм в почве, а также с содержанием Fe, Zn и Cu в самих травах при биологизации (табл. 4).
Полученные результаты свидетельствуют, что травы вовлекают в биологический круговорот больше Fe, Сu, Zn и Со, и меньше Mn по сравнению с их содержанием в почве. Относительный убывающий ряд содержания МЭ в травах имеет вид: Fe ˃ Zn ≥ Cu ˃ Mn ˃ Co. Накопление большого количества железа связано с участием этого элемента в синтетических процессах, содержание Mn в травах низкое, что отражает малую доступность его в щелочных почвах, а также антагонизм элементов в самом растении. Значительное накопление Zn и Cu в исследованных травах при их сочетании с МП может быть использовано для целей фиторемедиации загрязненных почв.
Исследование содержания МЭ в листьях груши сорта Таврическая показало, что на контроле было меньше Fe и Mn, чем в травах, причем содержание Fe, Сu и Zn в листьях находилось на уровне оптимума, Mn – было низким (Церлинг, 1990), а Co – высоким (табл. 5).
Таблица 5
Влияние биологизации на содержание МЭ (мг/кг) в листьях груши сорта Таврическая на подвое айва ВА 29 (2023 г.) (по Церлинг, 1990)
|
Вариант |
Контроль |
АБ |
МБК |
Среднее по фактору «задернение» |
|
Fe |
||||
|
ЕЗ |
86 |
93 |
93 |
90 |
|
СТ2 |
105 |
108 |
96 |
103* |
|
СТ4 |
136 |
134 |
92 |
121* |
|
Среднее по фактору «МП»; НСР05 = 1; НСР05ч.с. = 1 |
109 |
112* |
94* |
|
|
Mn |
||||
|
ЕЗ |
2,09 |
1,98 |
2,15 |
2,07 |
|
СТ2 |
1,86 |
2,29 |
1,90 |
2,02* |
|
СТ4 |
2,52 |
1,82 |
1,71 |
2,02* |
|
Среднее по фактору «МП»; НСР05 = 0,01; НСР05ч.с. = 0,02 |
2,16 |
2,03* |
1,92* |
|
|
Zn |
||||
|
ЕЗ |
32,1 |
72,2 |
57,2 |
53,8 |
|
СТ2 |
59,1 |
109,4 |
72,8 |
80,4* |
|
СТ4 |
70,7 |
91,8 |
36,9 |
66,5* |
|
Среднее по фактору «МП»; НСР05 = 1,9; НСР05ч.с. = 3,3 |
54,0 |
91,1* |
55,7 |
|
|
Cu |
||||
|
ЕЗ |
16,4 |
14,0 |
39,5 |
23,3 |
|
СТ2 |
23,2 |
28,5 |
30,4 |
27,3* |
|
СТ4 |
32,4 |
17,1 |
16,1 |
21,8* |
|
Среднее по фактору «МП»; НСР05 = 0,10; НСР05ч.с. = 0,17 |
24,0 |
19,9* |
28,7* |
|
|
Co |
||||
|
ЕЗ |
0,86 |
0,67 |
0,82 |
0,79 |
|
СТ2 |
0,89 |
1,15 |
1,18 |
1,07* |
|
СТ4 |
0,86 |
1,20 |
1,46 |
1,17* |
|
Среднее по фактору «МП»; НСР05 = 0,11; НСР05ч.с. = 0,19 |
0,87 |
1,00* |
1,15* |
|
Пояснение к таблице. АБ – азотобактерии; МБК – Микробиоком-Агро.
Применяемые приемы биологизации увеличивали содержание Fe, Zn и Co в листьях, причем на накопление Fe большее влияние оказывало задернение СТ4 – увеличивало на 31 мг/кг, при этом содержание элемента оставалось в пределах оптимума – 50–150 мг/кг (табл. 5).
Различные варианты опыта неоднозначно влияли на содержание Cu в листьях. При задернении почвы СТ2 и применении МБК как факторов содержание ее в листьях достоверно увеличивалось на 4 мг/кг, на вариантах СТ4 и АБ – снижалось на 1,5 и 4 мг/кг соответственно, однако ее содержание в листьях оставалось в пределах оптимального – 5−20 мг/кг (Церлинг, 1990). Все это происходило на фоне увеличения содержания доступной Cu в почве на этих вариантах и связано, скорее всего, со значительным накоплением Cu в травах на этих вариантах, что может защитить плодовое растение от избыточного накопления Cu при загрязнении почвы этим элементом.
Что касается Zn и Co, то их содержание в листьях груши увеличивалось значительно при совместном влиянии задернения и МП, особенно первого под действием сочетания СТ2 с АБ на 77 мг/кг, что может вызвать избыток этих элементов в растении и конкуренцию с Mn. Содержание Со в большей мере увеличивалось под действием сочетания СТ4 с МБК на 0,60 мг/кг или на 70 % от контроля по ЕЗ (табл. 5).
Значительное увеличение содержания Zn и Co в листьях груши вызывает существенное и достоверное снижение и так низкого содержания Mn в листьях, что связано с антагонизмом элементов в самом растении (табл. 5).
Связь между концентрацией Со и Mn в листьях при биологизации отрицательная сильная достоверная (r=−0,75). Визуальными признаками недостатка Mn для груши является наблюдаемый нами слабый хлороз, а также некроз молодых листьев.
Кроме того, часть элементов, имеющих низкое содержание в почве (Fe, Mn, Co) поглощается травами, что также может вызвать недостаток элементов в листьях. Так, применение МБК в среднем по фактору МП снижало содержание подвижного Mn в почве на 0,42 мг/кг по сравнению с контролем. В травах на этом же варианте произошло уменьшение содержания Mn на 0,31 мг/кг. В листьях груши в аналогичных условиях наблюдалось сокращение валового Mn на 0,24 мг/кг.
Установлено, что элементы по их концентрации в листьях груши расположились в следующий убывающий ряд: Fe ˃ Zn ˃ Cu ˃ Mn ˃ Co, что повторяет ряд, полученный для трав, но абсолютные величины были несколько иными.
Таким образом, на питание груши МЭ при биологизации могут влиять как содержание и изменение их концентрации в почве, поглощение травами, а также взаимосвязь элементов в самом плодовом растении. Избыточное накопление в почве Zn и Cu, а также Со в растении может вызвать недостаток Mn у плодового дерева при низком его содержании в карбонатных почвах с высоким содержанием органического вещества.
Расчеты накопления МЭ из почвы плодовым растением груши при биологизации агроценоза показывают, что все МЭ накапливались в листьях груши, кроме Mn, содержание которого в листьях было в 2−3 раза ниже, чем в почве (рис. 3). В контроле по ЕЗ Cu и Fe растением накапливаются в большом количестве, и степень этого увеличения возрастала в ряду ЕЗК < СТ2К < СТ4К (рис. 3). Поглощение Zn и Co было втрое ниже, чем первых двух элементов, однако содержание этих элементов в листьях было в 7–9 выше, чем в почве. Содержание Mn в листьях было ниже, чем в почве и практически не различалось по вариантам опыта (рис. 3).
МБК по ЕЗ способствовал значительному увеличению накопления грушей Cu (в 2,7 раза) и Fe (в 1,5 раза) по сравнению с контролем по ЕЗ. МП по фону ЕЗ увеличивали накопление Zn в 1,6–1,7 раза. Применение МП на фоне злаково-бобовых трав снижало накопление грушей Cu и Fe, особенно значительно на фоне СТ4 в 2,2–3,7 раза относительно СТ4К. При этом существенно увеличивалось поглощение Со в 1,8 раза, что, в свою очередь, снижало поглощение растением Mn (рис. 3).
В растении существует «взаимодействие» ионов – стимулирующее или ингибирующее действие увеличения концентрации одного иона на концентрацию другого (Эммерт, 1964; Фидлер, 1970). Ранее установлено, что ион Mn является антангонистом с ионами Fe, Zn, Cu и Со (Кабата-Пендиас, Пендиас, 1989). В нашем исследовании обнаружено увеличение последних ионов в листьях груши при биологизации, что может быть причиной низкого содержания Mn из-за снижения его поглощения растением в результате антагонизма с вышеперечисленными ионами. Для подтверждения данного явления мы рассчитали отношение Fe, Zn, Cu и Со (по отдельности) к Mn (рис. 4). Если это отношение увеличивается в вариантах с применением сеяных смесей трав и микробных препаратов, то можно считать причиной недостатка Mn в растении увеличение количества Fe, Zn, Cu и Со в листьях груши.
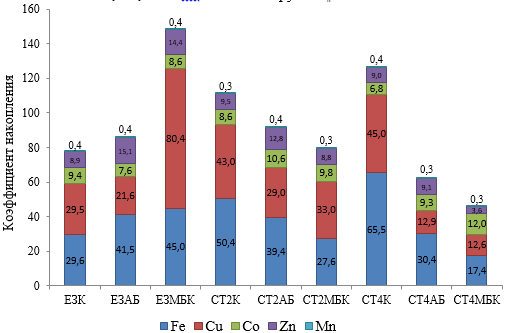
Рис. 3. Коэффициент накопления микроэлементов в листьях груши «Таврическая» при биологизации
Установлено, что отношение Zn и Mn было более чем втрое выше на вариантах с применением злаково-бобовых трав и АБ по сравнению с контролем по ЕЗ за счет значительного увеличения поглощения Zn. Отношение Fe/Mn было высоким во всех вариантах, но увеличение его по сравнению с контролем было самым большим на варианте СТ4 с АБ. Возможно, это связано со значительным повышением содержания доступного Fe для растения, обеспеченного активным штаммом-биоагентом этого препарата, что привело к увеличению отношения этих элементов в листьях груши.
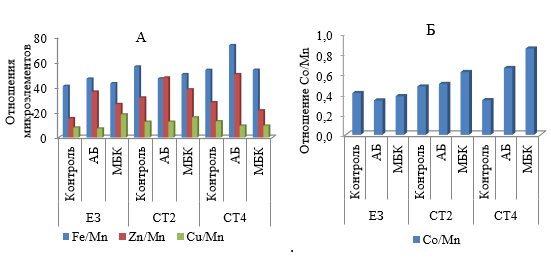
Рис. 4. Отношения микроэлементов в листьях груши сорта Таврическая при биологизации (2023 г.)
АБ – Азотобактерин 07-Агро; МБК – Микробиоком-Агро; ЕЗ – естественное задернение; СТ2 – смесь трав: Lolium multiflorum + Medicago sativa; СТ4 – смесь трав: L. multiflorum + M. sativa + Festuca pratensis + Trifolium pratense + Bromus inermis.
Что касается отношения Cu и Mn, то по фону ЕЗ и СТ2 оно значительно увеличивалось под влиянием МБК в 2,2 и 1,3 раза соответственно. По фону СТ4, наоборот, это соотношение снижалось под действием МП и было близким к контролю по ЕЗ (рис. 4 А). Отношение Co/Mn в листьях было меньше единицы и также увеличивалось под действием сочетаний сеяных трав с МП, максимально на варианте СТ4 с КМП (рис. 4 Б). Все это вызывает недостаток Mn в растении груши. Чтобы повысить содержание Mn в листьях до оптимального уровня (10–20 мг/кг) следует применять Mn-содержащие удобрения фолиарно (Минеев, 2006).
В целом биологизация агроценоза груши влияла на содержание МЭ в почве и растениях, а также поглощение и питание груши микроэлементами. В связи с установленным дефицитом доступного Mn в луговой карбонатной почве для груши, применение сочетания СТ2 с АБ создает более оптимальные условия для доступности растению МЭ, что не исключает удобрение груши Mn по листу для поддержания гомеостаза элемента в растении на оптимальном уровне.
ВЫВОДЫ
1. Установлено, что концентрация подвижных форм Fe, Mn и Co в почве была ниже регионального фона и по Mn и Co и была на уровне низкой обеспеченности для плодовых культур. Содержание подвижных Cu и Zn в почве было высоким, но не превышало ПДК и расположилось в следующий убывающий ряд: Mn ˃ Zn ˃ Fe ˃ Cu ˃ Co. Биологизация вызвала увеличение подвижности МЭ. Наибольшее влияние на содержание МЭ в почве оказывало задернение (доля влияния 30–60 % общей дисперсии), причем СТ4 был более эффективным из-за увеличения биоразнообразия трав при задернении. При этом МБК в сочетании с СТ4 максимально увеличивал содержание Fe и Zn, АБ – Mn и Co, содержание Сu увеличивали оба МП на том же фоне. Испытанные приемы биологизации, особенно МБК на фоне СТ4, способствовали некоторому относительно большему накоплению Zn по сравнению с Mn, что может вызвать недостаток Mn в растении груши.
2. Содержание валовых форм МЭ в травах было в основном оптимальным и высоким, Mn – низким. По содержанию в травах МЭ расположились в ряд Fe ˃ Cu ≥ Zn ˃ Mn ˃ Co. Недостаток Mn в травах также связан с дефицитом этого элемента в почве. В целом МП способствовали накоплению МЭ в биомассе сеяных трав за исключением Mn, содержание которого снижалось под действием МБК. Применение сеяных злаково-бобовых смесей трав с большой биомассой в сочетании с МП приводило к значительному накопление Zn на сочетании СТ2 с АБ в 2 раза и Cu – СТ4 с АБ в 4 раза по сравнению с контролем по ЕЗ. Такое накопление ТМ в травах может быть использовано для целей фиторемидиации загрязненных почв. В травах содержание Cu, Fe и Co зависело от содержания этих элементов в почве (r=0,46–0,67). Для Zn и Mn связи между концентрацией элементов в почве и листьях трав не обнаружено.
3. В листьях груши в контроле по ЕЗ содержание валовых форм МЭ было в основном оптимальным и высоким, Mn – низким. Ряд соотношения элементов был аналогичным, полученному для трав, но Zn в листьях было относительно больше, чем Сu. Применяемые приемы биологизации увеличивали содержание Fe, Zn и Co в листьях и снижали концентрацию Mn, содержание которого было значительно ниже оптимального для груши. Содержание Zn в варианте с задернением почвы СТ2 в сочетании с АБ увеличивалось втрое по сравнению с контролем по ЕЗ и становилось высоким, что может вызвать недостаток Mn и Fe в растении даже при достаточном их содержании в почве. Это проявилось во внешних симптомах недостаточности: слабый хлороз и некроз молодых листьев груши. Для улучшения питания груши Mn в условиях низкого содержания его подвижных форм в карбонатных почвах следует провести исследование по разработке оптимальных доз марганцевых удобрений, внесенных по листу для конкретных экологических условий.
4. Испытанные приемы влияли на биологическое поглощение МЭ растением груши, причем травы в большей степени стимулировали поглощение растением Fe и Сu. МП на фоне ЕЗ увеличивали поглощение Fe, МБК – Cu. На фоне злаково-бобовых трав МП, наоборот, снижали поглощение грушей этих элементов, в большей степени на фоне СТ4, что может свидетельствовать о более значительном поглощении этих элементов сеяными травами, особенно при увеличении их биоразнообразия по сравнению с естественным травяным покровом, а также самими бактериями, входящими в биопрепараты. При этом значительно увеличивалось поглощение грушей Со и снижалось поглощение Mn, что вызывает недостаток последнего в питании груши, как из-за его низкого содержания в почве, так и антагонизма с Fe, Zn, Cu и Со в самом растении.
Работа выполнена в рамках государственного задания Никитского ботанического сада по теме FNNS-2022-0005.
Акчурин А. Р., Костенко И. В. К оценке пригодности аллювиально-луговых почв Крыма под виноградники // Экологические проблемы садоводства и интродукции растений. Сборник научных трудов государственного Никитского ботанического сада. – 2008. – Т. 130. – С. 16–24.
Вернигорова Н. А., Колесников С. И., Казеев К. Ш. Оценка устойчивости почв и наземных экосистем Крыма к химическому загрязнению. Монография. – Ростов-на-Дону: Издательство Южного федерального университета, 2015. – 126 с.
Горбов С. Н., Безуглова О. С. Тяжелые металлы и радионуклеиды в почвах Ростовской агломерации. Монография. – Ростов-на-Дону – Таганрог: Изд-во Южного федерального университета, 2020. – 124 с.
Доспехов Б. А. Методика полевого опыта (с основами статистической обработки результатов исследований). – М.: Агропромиздат, 1985. – 351 с.
Иванова А. С. Микроэлементы в почвах под садами Крыма. – Ялта, 2002. – 36 с.
Ильин В. Б., Сысо А. И. Микроэлементы и тяжелые металлы в почвах и растениях Новосибирской области. – Новосибирск: Изд-во СО РАН, 2021. – 229 с.
Кабата-Пендиас А., Пендиас Х. Микроэлементы в почвах и растениях. – М.: Мир, 1989. – 439 с.
Казеев К. Ш., Колесников С. И., Акименко Ю. В., Даденко Е. В. Методы биодиагностики наземных экосистем: монография / [Отв. ред. К. Ш. Казеев]. – Ростов-на-Дону: Изд-во Южного федерального университета, 2016. – 356 с.
Карпухин А. И., Бушуев Н. Н. Влияние применения удобрений на содержание тяжелых металлов в почвах длительных полевых опытов // Агрохимия. – 2007. – № 5. – С. 76–84.
Клименко Н. И., Плугатарь Ю. В., Клименко О. Е. Создание полезных растительно-микробных взаимодействий в ризосфере декоративных растений. Монография. Симферополь: Ариал, 2019. – 110 с.
Клименко О. Е., Клименко Н. И., Дунаевская Е. В., Новицкая А. П., Новицкий М. Л. Влияние биологизации садового агроценоза на плодородие почвы, состояние и продуктивность персика // Агрохимический вестник. – 2020 – № 4. – С. 67–76. DOI: 10.24411/1029-2551-2020-10058
Клименко О. Е., Попов А. И. Биологизация интенсификационных процессов в агроценозе яблони (Malus domestica Borkh.) // Бюллетень государственного Никитского ботанического сада. – 2024. – Вып. 150. – С. 32–40. DOI:10.25684/0513-1634-2024-150-33-41
Кондаков А. К., Трунов Ю. В., Грезнев О. А., Сироткина О. А., Трунов А. А. Современная система минерального питания и удобрения плодовых и ягодных растений // Достижения науки и техники АПК. – 2009. – № 2. – С. 22–23.
Лисецкий Ф. Н., Зеленская Е. Я. Различия в содержании тяжелых металлов в почвах Южного берега Крыма (пространственно-временной анализ) // Экосистемы. – 2023. – Вып. 34. – С. 81–91.
Методические указания по определению тяжелых металлов в кормах и растениях и их подвижных соединений в почвах. − М.: ЦИНАО, 1993. – 40 с.
Минеев В. Г. Агрохимия. – М.: МГУ, Наука, 2006. – 720 с.
Плугатарь Ю. В., Клименко О. Е., Клименко Н. И., Сотник А. И., Орёл Т. И., Новицкий М. Л. Состав, свойства и рациональное использование почв садовых агроценозов долины р. Салгир (на примере отделения Никитского ботанического сада «Крымская опытная станция садоводства») // Сборник научных трудов государственного Никитского ботанического сада. – 2019. – Т. 148. – С. 5–21.
Скляр С. И., Валин Д. Н., Липиева Н. Н. Содержание подвижных форм микроэлементов в почвах республики Крым и его значение // Известия сельскохозяйственной науки Тавриды. – 2021. – № 25(188). – С. 5–18.
Соловьев Г. А. О методах определения азота, фосфора, калия, натрия, кальция, магния, железа и алюминия из одной навески после мокрого озоления // Проблемы почвоведения, агрохимии и мелиорации. – 1973. – С. 134–139.
Стойлов Г. П. Превращение форм соединений марганца при увлажнении и высыхании почв // Агрохимия. – 1967. – № 3. – С. – 92–97.
Трунов Ю. В., Трунова Л. Б. Достижения и проблемы российской науки в области минерального питания садовых растений // Плодоводство и виноградарство юга России. – 2013. – Вып. 23(5). – С. 121–130.
Фидлер В. Листовой анализ в плодоводстве. Перевод с немецкого Попеско И.Г., под редакцией и с предисловием Язвицкого М.Н. – М: Колос, 1970. – 56 с.
Церлинг В. В. Диагностика питания сельскохозяйственных культур. Справочник. – М.: Агропромиздат, 1990. – 235 с.
Эммерт Ф. Влияние взаимодействия ионов на состав растительных тканей // Анализ растений и проблемы удобрения. Перевод с английского Чебановой Т.Л., предисловие и редакция Петербургского А.В. – М.: Колос, 1964. – С. 218-233.
Sabir A. Improvement of grafting efficiency in hard grafting grape Berlandieri hybrid rootstocks by plant growth-promoting rhizobacteria (PGPR) // Scientia Horticulturae. – 2013. – Vol. 164, N 17. – P. 24–29.
Alvares-Tinaut M.C., Leal F., Recalde-Martinez L. R. Iron-manganese interaction and its relation to boron levels in tomato plants. – Plant and Soil. – 1980. – Vol. 55. – P. 377–388.
La Pera L., Dugo G., Rando R., Bella G. D., Maisaho R., Salvo F. Statistical study of the influence fungicide treatments (mancozeb, zoxamide and copper oxychloride) on heavy metal concentrations in Sicilian red wine // Food Additives and Contaminants. Part A Chemistry, Analysis, Control, Exposure and Risk Assessment. – 2008. – Vol. 25, N 3. – P. 302–313.
Vystavna, Y., Schmidt S., Klimenko O., Plugatar Yu., Klimenko N. I., Klimenko N. N. Species-dependent effect of cover cropping on trace elements and nutrients in vineyard soil and Vitis // Journal of the Science of Food and Agriculture. – 2020. – N 100. – P. 885–890.
