
УДК 582.477:581.151
Изменчивость биоморфологических
признаков шишек плосковеточника восточного (Platycladus orientalis (L.) Franco) в Крыму
Захаренко Г. С., Салогуб Р. В., Севастьянов В. Е.
Крымский федеральный университет имени В. И. Вернадского Симферополь, Республика Крым, Россия
Изучена внутривидовая изменчивости шишек у плосковеточника восточного (Platycladus orientalis (L.) Franco) в декоративных насаждениях в Предгорном Крыму и на ЮБК, а также на четырёх участках лесных культур в приморской зоне Евпатории. Определена эндогенная, индивидуальная и междемовая изменчивость структурных признаков шишек: длины и толщины во взаимно перпендикулярных направлениях в наиболее толстой части, числа чешуй и семян, длины апофиза наиболее развитых чешуй. На индивидуальном уровне среднестатистические значения параметров шишек составляют: длина – 13,8–22,6 мм (6 % ≤ С ≤ 17 %), больший диаметр – 12,0–18,6 мм (7 % ≤ С ≤ 19 %), меньший диаметр – 12,0–15,7 мм, число чешуй – 5,9–7,6 шт. (7 % ≤ С ≤ 14 %), длина апофиза – 0,16-2,09 мм (29 % ≤ С ≤ 106 %), число семян – 4,5–7,3 шт. (9 % ≤ С ≤ 36 %). Полученные результаты подтверждают данные об органоспецифичности уровней изменчивости структурных элементов генеративных органов у голосеменных растений. Широкая амплитуда изменчивости длины апофиза связана с его атавистической природой вследствие морфологического проявления кроющей чешуи, сросшейся с семенной чешуёй на ранних этапах эволюции современных видов семейства Cupressaceae, и кондупликатным развитием чешуй их шишек.
У плосковеточника, как и в природных популяциях сосны обыкновенной и ели европейской, уровни эндогенной и индивидуальной изменчивости совпадают. Это указывает на сбалансированность фенетического полиморфизма крымской интродукционной популяции Platycladus orientalis и свидетельствует о высоком микроэволюционном потенциале вида в условиях изоляции и возможности формирования в южных районах России в течение 20–25 перекрывающихся семенных поколений местной репродукции интродукционных популяций, сопоставимых по уровню фенотипической изменчивости природным популяциям голосеменных древесных растений.
Ключевые слова: плосковеточник, изменчивость, морфологические признаки, шишки, интродукция.
ВВЕДЕНИЕ
Плосковеточник восточный является представителем монотипного рода Рlatycladus Spach семейства Cupressaceae Bartlett. Его родиной считается Китай (Farjon, Filer, 2013), при том что самовоспроизводящиеся популяции плосковеточника встречаются от Ирана до Кореи на высотах от 300 до 3300 м над уровнем моря (Fu еt al., 1999). Коренных лесов с участием плосковеточника восточного практически не сохранилась. Сегодня чаще всего данный вид можно видеть в составе вторичных (производных) лесных экосистем, глубоко преобразованных в результате деятельности человека.
В природных условиях деревья плосковеточника способны доживать до 1000 лет, достигая более 20 м в высоту и до 2 м в диаметре ствола.
В течение многих веков плосковеточник используется при создании фитомелиоративных и декоративных насаждений на территории Китая и южных районов Центральной Азии. В результате интродукционной деятельности культурный ареал рассматриваемого вида охватывает все континенты Земли (Guoqing, 2016). В декоративных насаждениях Пекина насчитывается большое число многовековых деревьев плосковеточника, взятых под охрану государства (Sun еt al., 1992). Лесоводы Китая рассматривают плосковеточник как важную лесомелиоративную породу для закрепления склонов (Wang еt al., 2013). Изучаются вопросы семенного размножения и выращивания посадочного материала данного вида (Yang еt al., 2011; Liu еt al., 2013).
ISSN 2414-4738 Published by V. I. Vernadsky Crimean Federal University, Simferopol
В Крыму плосковеточник введен в культуру в 1813 году (Забелин, 1939). Благодаря засухоустойчивости, невысокой требовательности к плодородию почв, относительной морозостойкости и декоративным качествам, он получил широкое распространение в зеленом строительстве и степном лесоразведении в Крыму, степных районах Северного Кавказа и Северного Причерноморья.
Для плосковеточника в условиях культуры в Крыму характерно очень быстрое вступление растений семенного происхождения в репродуктивную фазу. По нашим наблюдениям, в благоприятных условиях выращивания в питомнике первые женские шишки закладываются у растений семенного происхождения в возрасте четырёх лет. Исходя из этого, можно полагать, что в культуре в Крыму этот вид представлен многими поколениями местной семенной репродукции, что позволяет изучать этот вид с позиций популяционной биологии (Некрасов, 1980; Яблоков, 1987).
Изучение особенностей репродуктивного развития выявило эндогенную и индивидуальную изменчивость ритмов развития, обилия закладки микростробилов и женских шишек, процесса опыления и диссеминации у плосковеточника во всех районах Крыма, а также значительную вариабельность посевных качеств семян как в урожае отдельно взятого дерева, так и у рядом растущих деревьев (Захаренко и др., 2022а, 2022б).
К настоящему времени изучение внутривидовой изменчивости морфологических признаков у плосковеточника восточного в условия интродукции в нашей стране и за ее пределами ограничивалось отбором и вегетативным размножением форм, отличающихся габитусом кроны, ветвлением, окраской листьев и представляющих интерес для декоративного садоводства (Krüssmann, 1985). Изменчивость морфологических признаков репродуктивных органов, часто используемая в фенетике древесных растений (Мамаев, 1973), у данного вида практически не изучена. Приводятся лишь краткие сведения об общем виде и размерах зрелых шишек и микростробилов плосковеточника (Fu еt al., 1999).
Устойчивость вида и его способность приспосабливаться к изменяющимся условиям среды при интродукции во многом зависит от генетического разнообразия совокупности растений, имеющихся в районе культуры. Одним из важных направлений изучения генетического разнообразия вида является оценка его фенотипической изменчивости по возможно большему числу признаков как в природном ареале, так и в условиях интродукции (Яблоков, 1987; Яблоков, Ларина, 1985). Большой интерес такие исследования представляют для оценки адаптивного потенциала и перспектив использования хозяйственно ценных иноземных древесных растений в декоративном садоводстве и лесоразведении. В связи со сложностью изучения адаптивных наследственных признаков, имеющих полигенную природу, в качестве первого шага оценки генетического полиморфизма популяций используют фенетический анализ изменчивости морфологических признаков вегетативных и генеративных признаков. При этом особый интерес представляют признаки, характеризующиеся минимальной вариабельностью на уровне отдельного дерева, которая характеризуется коэффициентом их вариации.
Накопление данных об изменениях в фенофонде открывает возможность проследить тренд микроэволюционных сдвигов и выявить особенности формирования интродукционных популяций в череде семенных поколений местной репродукции. Это также дает возможность управлять микроэволюционным процессом методами селекции. Такие исследования заслуживают особого внимания в условиях возможного глобального изменения климата, когда в результате смещения границ природно-климатических зон основой ведения лесного хозяйства могут стать иноземные древесные растения, к числу которых в засушливых условиях юга России относится плосковеточник восточный.
Целью проведенных нами исследований было изучение изменчивости морфологических признаков шишек плосковеточника восточного в Крыму.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Объектом исследования служили шишки деревьев плосковеточника в декоративных насаждениях Южного берега и Предгорной зоны Крыма, а также на участках лесных культур в Евпаторийском лесничестве. Шишки собирали непосредственно перед началом их раскрывания. Для биометрического анализа с каждого из 36 деревьев собирали по 25 шишек в средней части кроны.
У каждой шишки с помощью штангенциркуля с точностью 0,1 мм измеряли длину тела
(L) и толщину в двух взаимно перпендикулярных направлениях в её наиболее толстой части (D – диаметр максимальный и d – диаметр минимальный), а также расстояние между кончиками шипов наиболее развитой, как правило, второй нижней пары чешуй (B). Длину выростов чешуй (k) определяли расчётным путём по формуле: k=(B – D)/2.
Для сравнительного визуального анализа внешних признаков шишек отдельно взятых деревьев измерение сопровождалось их фотографированием. Каждая измеренная шиша помещалась в отдельный бумажный пакет. После высыхания шишки подсчитывали число чешуй и развившихся в ней семян.
Полученный числовой материал обработан с помощью пакета прикладных программ статистического анализа Excel 2016. Уровень изменчивости показателей оценивали по шкале, предложенной С. А. Мамаевым (1973).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ОБСУЖДЕНИЕ
У плосковеточника восточного шишки закладываются на концах побегов в периферийной части кроны часто в виде гроздей. В таких гроздях шишки, расположенные на концах наиболее развитых боковых побегов, имеют большие размеры, чем развивающиеся на ниже расположенных на материнской ветви побегах.
Шишки образованы накрест супротивно расположенными парами чешуй, нижние из которых остаются недоразвитыми. В разной степени недоразвитыми или сросшимися в шишках отдельных деревьев также остаются чешуи терминальной пары.
Закладка и развитие отдельно кроющих и семенных чешуй у плосковеточника восточного не наблюдается, то есть шишка образована, по определению А. Л. Тахтаджяна, из
«простых» чешуй (Тахтаджян, 1956).
В поперечном сечении, проведенном через предусмотренные нашей методикой точки измерения толщины шишки в её наиболее утолщённой части, её тело по форме близко к овальной. Больший диаметр сечения тела шишки (D) совпадает с серединой наиболее развитой пары чешуй. Тело шишки без учёта выростов чешуй по форме варьирует от сплюснутой с боков шаровидной до овоидной.
Визуальный анализ шишек у плосковеточника восточного, собранных в декоративных и лесомелиоративных насаждениях Крыма, позволяет видеть изменчивость их формы и размеров как на индивидуальном, так и эндогенном уровнях (рис. 1). Одним из существенных морфологических признаков, характеризующих индивидуальную изменчивость деревьев по морфологическим признакам шишек, является степень развития у чешуй выростов (апофизов). На индивидуальном уровне форма апофиза варьирует от небольших чешуевидных (рис. 1 Г, Ж) до крупных клювовидных выростов (рис. 1 А, Д). У шишек отдельных деревьев шиповидный или клювовидный вырост бывает более сильно развит только на верхних чешуях (рис. 1 Б, В, Е). Шишки с апофизами, наиболее развитыми у верхних пар чешуй (рис. 1, Б), приобретают бокаловидную форму.
Сочетание таких морфологических признаков как длина и толщина тела шишки в двух взаимно перпендикулярных направлениях, форма и размеры выростов (апофизов) чешуй и их приуроченность к определённым парам чешуй делают эти морфологические признаки четкими показателями индивидуальных особенностей деревьев плосковеточника восточного и могут рассматриваться в качестве фенов.
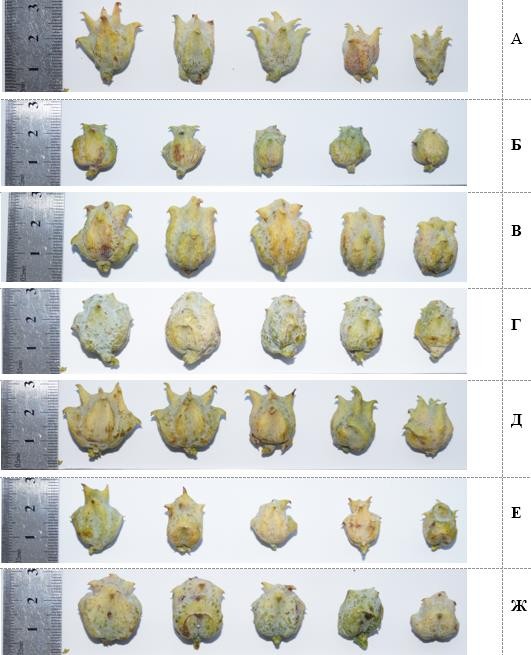
Рис. 1. Индивидуальная и эндогенная морфологическая изменчивость шишек плосковеточника восточного в Крыму
А–Ж – образцы шишек отдельных деревьев.
Результаты изучения эндогенной изменчивости зрелых шишек у отдельных деревьев плосковеточника, представленные в таблицах 1 и 2, показывают, что различия между шишками в урожае отдельно взятого дерева по длине тела составляет от 3,8 мм (деревья № 1/1
и 3/4) до 12,1 мм (дерево №2/3), при среднем значении этого показателя соответственно 19,7 мм и 12,2 мм в первом случае, и 17,8 мм во втором. Эндогенное варьирование данного показателя у большинства деревьев во всех местах произрастания характеризуется очень низким или низким уровнями изменчивости (С÷5–12 %), и лишь у одного из исследуемых деревьев составляет 17 %, то есть оценивается средним уровнем по шкале С. А. Мамаева.
Таблица 1
Эндогенная изменчивость морфологических признаков тела шишек плосковеточника восточного в Крыму
| № дерева* | Морфологические признаки шишек | ||||||||
| Длина тела шишки (H), мм | Больший диаметр сечения тела
шишки (D), мм |
Меньший диаметр сечения
тела шишки (d), мм |
|||||||
| L±m | lim L | С,
% |
D±m | lim D | С,
% |
d±m | lim d | С,
% |
|
| 1/1 | 19,7±0,19 | 17,7–21,5 | 5 | 14,5±0,41 | 12,7–25,5 | 15 | 13,1±0,46 | 11,0–25,3 | 19 |
| 1/3 | 21,3±0,38 | 18,0–25,8 | 10 | 17,0±0,44 | 13,0–25,6 | 14 | 15,7±0,39 | 12,7–24,6 | 14 |
| 1/6 | 15,5±0,27 | 12,5–19,5 | 10 | 13,3±0,25 | 11,2–17,0 | 10 | 12,0±0,18 | 10,0–14,2 | 8 |
| 2/1 | 20,8±0,25 | 17,7–23,0 | 7 | 17,3±0,23 | 15,0–19,4 | 7 | 14,4±0,19 | 12,3–16,7 | 7 |
| 2/3 | 17,8±0,56 | 11,5–23,6 | 17 | 13,5±0,32 | 9,8–16,0 | 12 | 12,8±0,30 | 9,8–16,0 | 13 |
| 2/7 | 19,7±0,36 | 16,1–24,2 | 8 | 14,0±0,17 | 12,2–16,3 | 7 | 12,9±0,15 | 10,9–15,0 | 6 |
| 3/1 | 20,5±0,30 | 16,0–24,4 | 9 | 15,9±0,31 | 12,5–19,7 | 12 | 14,1±0,20 | 10,6–17,1 | 9 |
| 3/2 | 17,2±0,22 | 13,8–19,6 | 8 | 17,0±0,22 | 14,5–20,0 | 8 | 14,9±0,19 | 11,7–17,3 | 8 |
| 3/4 | 12,2±0,11 | 10,3–14,1 | 6 | 12,0±0,14 | 9,8–13,9 | 7 | 9,8±0,11 | 8,0–11,6 | 7 |
| 4/1 | 19,1±0,25 | 15,2–22,2 | 8 | 16,5±0,10 | 11,3–19,9 | 11 | 14,6±0,20 | 12,3–17,4 | 9 |
| 4/2 | 13,8±0,21 | 10,8–16,4 | 9 | 12,7±0,17 | 10,3–14,8 | 8 | 10,6±0,15 | 9,0–12,6 | 9 |
| 4/4 | 15,9±0,23 | 12,3–18,5 | 9 | 14,7±0,27 | 12,2–18,7 | 12 | 12,9±0,24 | 10,6–17,3 | 12 |
| 5/1 | 18,4±0,27 | 14,7–21,7 | 9 | 17,1±0,27 | 13,0–20,6 | 10 | 17,1±0,27 | 11,7–17,6 | 10 |
| 5/3 | 16,2±0,32 | 12,2–19,5 | 12 | 17,1±0,33 | 11,7–22,0 | 12 | 14,2±0,30 | 10,8–19,1 | 13 |
| 5/4 | 17,8±0,25 | 13,4–20,5 | 12 | 13,9±0,23 | 12,1–18,7 | 11 | 12,6±0,18 | 10,0–16,0 | 9 |
| 6/1 | 15,4±0,26 | 11,0–18,5 | 11 | 13,7±0,26 | 9,9–18,9 | 12 | 11,7±0,20 | 8,6–15,8 | 11 |
| 6/3 | 22,6±0,23 | 19,1–25,5 | 7 | 18,6±0,36 | 14,2–23,9 | 12 | 15,5±0,20 | 12,6–17,9 | 8 |
| 6/4 | 18,3±0,30 | 13,6–22,2 | 10 | 13,7±0,29 | 10,0–18,9 | 13 | 12,4±0,25 | 13,6–22,2 | 10 |
Причиной относительно широкого варьирования длины тела шишки, когда имеет место двукратное превышение рассматриваемого показателя (дерево № 2/3), очевидно является высокий урожай шишек у дерева. При этом развитие более крупных шишек на побегах, сформированных в периферической части кроны, ближе к концам ветвей, вероятно, связано с особенностями перераспределения пластических веществ между побегами различных порядков в системе ветвления относительно вершины осевого побега материнской ветви.
Связь между размерами шишек, обилием их закладки и положение на побегах различных порядков ветвления в пределах ветви была обнаружена нами также у кипарисов (Захаренко, 2006). Описанное у многих растений различие плодовых образований по размерам при их массовой закладке, рассматривается как результат конкурентных отношений между ними (Леопольд, 1968).
Аналогичная картина эндогенной изменчивости наблюдается по показателям диаметра шишки D и d. Различия между максимальным и минимальным значениями этих показателей также могут достигать двукратных значений. Однако причины этих различий во многом иные, чем по длине тела шишки, и обусловлены особенностями формы отдельных шишек, отличающихся количеством пар чешуй и степенью развития апофиза (табл. 2).
Шишки по количеству слагающих их чешуй варьируют в узких пределах, поскольку у абсолютного большинства деревьев их число не превышает 8 штук (4 пары). На эндогенном
Таблица 2
Эндогенная изменчивость размеров апофиза и счётных морфологических признаков шишек плосковеточника восточного в Крыму
| № дерева* | Морфологические признаки шишек | ||||||||
| Длина апофиза (k), мм | Число чешуй в шишке
(n), шт. |
Число семян в шишке (N),
шт. |
|||||||
| k±m | lim k | С,
% |
n ±m | lim n | С,
% |
N±m | lim N | С,
% |
|
| 1/1 | 1,33±0,07 | 0,55–2,20 | 30 | 7,0±0,18 | 6–8 | 14 | 7,0±0,30 | 4–10 | 23 |
| 1/3 | 1,35±0,11 | 0,35–2,40 | 43 | 6,3±0,16 | 4–8 | 14 | 6,1±0,26 | 2–10 | 23 |
| 1/6 | 0,64±0,07 | 0,01–1,65 | 62 | 6,1±0,13 | 4–8 | 12 | 5,4±0,21 | 4–9 | 20 |
| 2/1 | 2,09±0,12 | 0,70–2,40 | 28 | 6,0±0,03 | 6–7 | 3 | 6,0±0,01 | 4–8 | 17 |
| 2/3 | 1,57±0,12 | 0,40–,45 | 43 | 7,1±0,18 | 6–8 | 14 | 7,3±0,30 | 5–10 | 21 |
| 2/7 | 0,66±0,09 | 0,01–1,75 | 73 | 5,9±0,07 | 4–6 | 7 | 5,9±0,10 | 5–7 | 9 |
| 3/1 | 0,65±0,09 | 0,01–1,70 | 89 | 6,9±0,15 | 6–8 | 13 | 6,8±0,20 | 5–10 | 18 |
| 3/2 | 1,51±0,12 | 0,01–3,40 | 50 | 6,8±0,15 | 6–8 | 14 | 7,1±0,25 | 5–10 | 22 |
| 3/4 | 1,43±0,06 | 0,01–2,25 | 29 | 6,1±0,06 | 6–8 | 7 | 6,0±0,12 | 5–9 | 12 |
| 4/1 | 1,09±0,07 | 0,03–2,25 | 42 | 6,5±0,13 | 6–8 | 13 | 6,0±0,14 | 4–10 | 14 |
| 4/2 | 0,41±0,06 | 0,01–1,20 | 94 | 6,1±0,08 | 4–8 | 9 | 5,1±0,16 | 3–7 | 20 |
| 4/4 | 1,06±0,10 | 0,02–2,25 | 61 | 6,2±0,12 | 4–8 | 12 | 5,0±0,21 | 2–10 | 26 |
| 5/1 | 1,09±0,07 | 0,02–2,25 | 42 | 6,8±0,14 | 6–8 | 13 | 5,1±0,20 | 2–8 | 25 |
| 5/3 | 1,12±0,08 | 0,01–2,40 | 46 | 6,7±0,14 | 5–8 | 14 | 5,3±0,27 | 2–9 | 32 |
| 5/4 | 1,06±0,11 | 0,02–2,25 | 61 | 6,2±0,08 | 6–8 | 8 | 5,6±0,15 | 4–8 | 17 |
| 6/1 | 0,08±0,04 | 0,00–1,05 | 99 | 6,1±0,09 | 5–8 | 9 | 4,7±0,22 | 2–8 | 30 |
| 6/3 | 0,16±0,06 | 0,01–1,50 | 106 | 7,6±0,13 | 6–8 | 11 | 7,4±0,29 | 4–10 | 24 |
| 6/4 | 1,75±0,13 | 0,02–3,35 | 48 | 6,2±0,08 | 5–8 | 9 | 4,5±0,25 | 1–7 | 36 |
Примечание к таблице. * – №№ деревьев: 1/1, 1/3, 1/6 – Южный берег Крыма; 2/1, 2/3, 2/7 – Предгорная зона Крыма; 3/1, 3/2, 3/4 – участок 1, Евпаторийское лесничество; 4/1, 4/2, 4/4 – участок 2, Евпаторийское лесничество; 5/1, 5/3, 5/4 – участок 3, Евпаторийское лесничество; 6/1, 6/3, 6/4 – участок 4, Евпаторийское лесничество.
уровне их число варьирует от 4–6 до 4–8 штук. Такое варьирование обусловлено максимально и минимально возможным числом пар чешуй в шишке, которое не может быть меньше двух и максимально четырёх пар чешуй.
Эндогенная изменчивость рассмотренных мерных признаков и количества чешуй в шишках у большинства деревьев характеризуется низким уровнем изменчивости (С ≤ 12 %), и лишь в выборках единичных деревьев достигает среднего уровня (С ≤ 20 %). Среди рассматриваемых морфологических признаков шишек плосковеточника восточного наиболее вариабельным является длина апофиза (k). Эндогенная изменчивость этого признака характеризуется уровнями изменчивости от повышенного (С÷20–30 %) у деревьев № № 1/1, 2/1, 3/4 до очень высокого (С>40 %) у остальных. Как видно из таблиц 1 и 2, повышенный уровень эндогенной изменчивости характерен для деревьев с достаточно крупными апофизами чешуй, а очень высокий – со среднестатистическим значением его длины менее 1 мм. При визуальном описании шишек форму таких коротких апофизов можно относить к чешуям или коротким шипам.
Обнаруженная нами большая амплитуда эндогенной изменчивости длины апофиза характерна и для других хвойных. Например, у сосны обыкновенной на Урале этот признак также характеризуется высоким уровнем эндогенной изменчивости – С÷24–39 %. (Мамаев, 1973).
В процессе формирования шишки на каждой чешуе развиваются две, а на чешуях верхней нормально развитой пары часто одна семяпочка (Jagel, Dӧrken, 2015). При наличии в шишке 4–8 чешуй в шишке потенциально могут развиться от 8 до 16 семян. Однако число резвившихся семян в отдельно взятой шишке определяется не только потенциально
возможным количеством чешуй и заложенных на них семяпочек, но и, как показали ранее проведенные нами исследования, у каждого дерева это определяется также его индивидуальными феноритмическими особенностями и сочетанием погодно-климатических факторов в период формирования генеративных органов и опыления (Захаренко и др., 2022б). В результате этого наблюдается более высокий уровень эндогенной изменчивости числа семян в шишках относительно аналогичного показателя, характеризующего изменчивость числа чешуй в них. Если коэффициент эндогенной изменчивости числа чешуй С÷3–14 %, то числа семян С÷9–36 %.
Приведенные в таблицах 1 и 2 и на рисунке 1 данные показывают, что у плосковеточника восточного наблюдается большое сходство деревьев по уровню эндогенной изменчивости биометрических показателей шишек, при достаточно чётком различии отдельных деревьев по форме шишек.
Данные по эндогенной изменчивости мерных и счётных признаков шишек плосковеточника восточного в насаждениях (демах), созданных в различных районах Крыма, делают возможным оценить взаимосвязь между изученными признаками. Расчёт коэффициентов корреляции показал, что математически достоверная связь существует как между мерными, так и счётными признаками (табл. 3). Наиболее взаимозависимыми признаками являются размеры тела и число морфологически нормально развитых семян в шишке. Недостаточно высокая частота достоверной корреляции между длиной шишки и числом семян у деревьев в пределах рассмотренной совокупности деревьев позволяет лишь условно говорить о независимости процессов формирования морфологических признаков шишки от числа сформировавшихся в ней семян, поскольку не учтено число развивавшихся и отмерших семян на начальных этапах эмбриогенеза.
Таблица 3
Доля деревьев плосковеточника восточного в Крыму с достоверной корреляционной связью (r≥0,55) между морфологическими признаками шишек, %
| Морфометрические признаки шишек | Длина шишки | Диаметр максималь– ный | Диаметр минимальный | Число чешуй в шишке | Число семян в шишке |
| Диаметр максимальный | 55 | ||||
| Диаметр минимальный | 76 | 79 | |||
| Число чешуй в шишке | 21 | 39 | 39 | ||
| Число семян в шишке | 45 | 70 | 52 | 55 | |
| Длина апофиза | 0 | 4 | 9 | 21 | 9 |
Различия в корреляции между структурными признаками шишки, вероятно, связаны с определённой автономией векторов развития морфологических элементов шишки в пределах общего морфогенетического пространства, задаваемого генотипом дерева с учётом влияния комплекса внешних и внутренних физиологических факторов.
Оценка эндогенной и межгрупповой изменчивости показало, что наиболее вариабельными признаками, как в урожае шишек отдельно взятого дерева, так и в выборках деревьев из разных районов Крыма, являются число семян в шишке и размеры апофиза (табл. 4).
Вариабельность числа морфологически нормально сформированных семян и их качество, зависит не только от индивидуальных особенностей дерева, но и, как показали наши исследования (Захаренко и др., 2022а, 2022б), условий опыления конкретного дерева. На это указывает и тот факт, что в шишках, образованных из четырёх пар чешуй, при потенциально возможном развитии 16 семян, их максимальное число в одной шишке у деревьев во всех районах Крыма не превышало 10 штук.
Таблица 4
Междемовая изменчивость морфологических признаков шишек плосковеточника восточного в Крыму
| Морфологические признаки шишек | |||||||||||
| Длина, мм | Наибольший
диаметр, мм |
Наименьший
диаметр, мм |
Число чешуй,
шт. |
Число семян,
шт. |
Длина апофиза,
мм |
||||||
| L±m | С,
% |
D±m | С,
% |
d±m | С,
% |
n±m | С,
% |
N±m | С,
% |
k±m | С,
% |
| Южный берег Крыма | |||||||||||
| 19,9±0,21 | 14 | 14,9±0,16 | 15 | 13,0±0,16 | 17 | 6,4±0,06 | 13 | 5,9±0,12 | 27 | 1,27±0,05 | 54 |
| Предгорная зона Крыма | |||||||||||
| 19,1±0,18 | 14 | 18,1±0,17 | 12 | 14,8±0,13 | 13 | 6,5±0,06 | 14 | 6,6±0,09 | 20 | 1,65±0,05 | 41 |
| Участок 1, Евпаторийское лесничество | |||||||||||
| 16,8±0,22 | 18 | 14,5±0,16 | 16 | 12,4±0,15 | 17 | 6,8±0,06 | 14 | 7,0±0,11 | 14 | 0,87±0,05 | 87 |
| Участок 2, Евпаторийское лесничество | |||||||||||
| 17,1±0,17 | 14 | 14,7±0,13 | 13 | 13,0±0,13 | 14 | 6,4±0,06 | 13 | 5,6±0,08 | 20 | 0,09±0,04 | 99 |
| Участок 3, Евпаторийское лесничество | |||||||||||
| 18,3±0,22 | 17 | 15,5±0,18 | 17 | 13,4±0,14 | 15 | 6,6±0,07 | 14 | 5,6±0,14 | 37 | 1,28±0,09 | 107 |
| Участок 4, Евпаторийское лесничество | |||||||||||
| 17,3±0,13 | 11 | 16,0±0,15 | 13 | 13,7±0,13 | 13 | 6,7±0,06 | 14 | 5,7±0,10 | 24 | 0,65±0,07 | 143 |
Изменчивость длины апофиза, вероятно, зависит от комплекса физиологических и морфогенетических процессов внутри дерева, которые определяют особенности морфологического развития опылённых шишек как целостных органов и морфогенеза составляющих их структурных элементов. Наибольшей вариабельностью по длине апофиза на эндогенном уровне характеризовались деревья с обильным урожаем и гроздеобразным размещением шишек.
Известно, что у представителей семейства кипарисовые чешуи шишек имеют две системы проводящих пучков, что является следствием срастания семенной и кроющей чешуй на ранних этапах филогенеза представителей этого семейства (Hagerup, 1933; Lemoine- Sebastian, 1968). Можно полагать, что наличие апофиза является атавизмом, филогенетически обусловленным кондупликатной природой формирования простых чешуй шишки. С этой точки зрения, апофиз является морфологическим проявлением кроющей чешуи, утратившей в филогенезе вида морфологическую самостоятельность. По-видимому, этим объясняется отсутствие корреляции между размерами апофиза и остальными счётными и мерными признаками шишек.
Полученные нами данные об изменчивости шишек у плосковеточника восточного позволяют считать, что у данного вида, как и у сосен и елей в природных популяциях, уровень эндогенной изменчивости признаков, характеризующих размеры шишек и число чешуй в них, органоспецифичен и обусловлен видовыми особенностями морфо- и органогенеза (Мамаев, 1973).
Оценить силу влияния индивидуальных особенностей деревьев и условий произрастания на варьирование морфометрических признаков шишек на статистическом уровне позволяет двухфакторный дисперсионный анализ. При этом отметим, что влияние фактора «условия произрастания» включает в себя не только собственно почвенно-климатические условия произрастания, но и не учтённое в данном случае обилие урожая шишек у деревьев, то есть особенности перераспределения у деревьев пластических веществ в системе ветвления и между шишками, особенно при их обилии и гроздеобразном размещении.
В результате двухфакторного дисперсионного анализа обнаружено (табл. 5), что наиболее сильное влияние индивидуальных особенностей деревьев сказывается на изменчивости таких показателей размера шишки как максимальный и минимальный диаметр (соответственно 13,1 % и 16,5 %). Влияние же индивидуальных особенностей дерева на
изменчивость длины шишки среди мерных и остальных признаков оказалось минимальным – 3,8 %, в то время как сила влияния условий произрастания наибольшая – 20,9 %. На сложную природу влияния взаимодействия индивидуальных особенностей и условий произрастания на изменчивость мерных признаков указывает величина его силы, составляющая около 34 % на размеры тела шишки и более 41 % на длину апофиза.
Таблица 5
Результаты двухфакторного дисперсионного анализа морфометрических показателей шишек плосковеточника восточного в Крыму
| Признаки шишек | Факторы | Критерий
Фишера* |
Сила влияния
фактора (h2), % |
| Максимальный диаметр | Индивидуальные особенности деревьев | 47,7 | 13,1 |
| Условия произрастания | 14,1 | 3,9 | |
| Взаимодействие | 30,6 | 33,5 | |
| Минимальный диаметр шишки | Индивидуальные особенности деревьев | 68,7 | 16,5 |
| Условия произрастания | 21,0 | 5,1 | |
| Взаимодействие | 36,2 | 34,8 | |
| Длина шишки | Индивидуальные особенности деревьев | 16,9 | 3,8 |
| Условия произрастания | 91,8 | 20,9 | |
| Взаимодействие | 37,3 | 34,0 | |
| Число чешуй в шишке | Индивидуальные особенности деревьев | 21,4 | 9,2 |
| Условия произрастания | 3,8 | 1,6 | |
| Взаимодействие | 6,6 | 11,3 | |
| Число семян в шишке | Индивидуальные особенности деревьев | 13,2 | 5,0 |
| Условия произрастания | 29,3 | 11,2 | |
| Взаимодействие | 9,7 | 14,6 | |
| Длина апофиза | Индивидуальные особенности деревьев | 28,7 | 6,8 |
| Условия произрастания | 37,7 | 8,9 | |
| Взаимодействие | 44,1 | 41,6 |
Примечание к таблице. * – Во всех случаях Fфакт.> F0,0001.
Влияние индивидуальных особенностей деревьев, условий произрастания и их взаимодействие на изменчивость числа чешуй и семян невелика и суммарно составляет менее 30 %, то есть развитие этих признаков шишек регулируется более сложным комплексом физиолого-биохимических и морфогенетических факторов, складывающихся в дереве как относительно целостной системе частей и органов различной степени соподчинённости.
ЗАКЛЮЧЕНИЕ
Результаты исследований эндогенной и индивидуальной изменчивости шишек показали, что у плосковеточника восточного в условиях культуры в Крыму уровни изменчивости рассмотренных признаков совпадают с данными аналогичных исследований природных популяций елей и сосен и подтверждают вывод об органоспецифичности уровней изменчивости структурных элементов генеративных органов у голосеменных растений.
Корреляция между структурными признаками шишки в значительной мере определяется числом развивающихся в шишке семян, а также индивидуальными векторами их морфогенеза в рамках общего для вида морфогенетического пространства. Отсутствие корреляции между размерами апофиза и другими признаками шишки указывает на его атавистическую природу, филогенетически обусловленную кондупликатной природой формирования простых чешуй шишки у представителей семейства Cupressaceae.
Отсутствие различий эндогенного и индивидуального уровней изменчивости одноименных признаков позволяет с большой вероятностью считать, что в Крыму в череде поколений местной семенной репродукции сформировалась интродукционная популяция плосковеточника восточного со сбалансированным фенетическим полиморфизмом по
структурным признакам шишек. Это свидетельствует о высоких темпах микроэволюции плосковеточника восточного в условиях изоляции и широкой культуры, а также о возможности формирования на юге России в течение 20–25 перекрывающихся семенных поколений местной репродукции интродукционных популяций, сопоставимых по уровню фенотипической изменчивости природным популяциям голосеменных древесных растений.
Список литературы
Атрохин В. Г. Biota orientalis (L.) Endl. // Древесные породы мира. Т. 3. Древесные породы СССР / [Под ред.
К. К. Калуцкого]. – М.: Лесная промышленность, 1982. – С. 212–213.
Забелин И. А. Gymnospermae – голосеменные // Труды Никитcкого ботанического сада. – 1939. – Т. 22. – Вып. 1. – С. 35–178.
Захаренко Г. С. Биологические основы интродукции и культуры видов рода кипарис (Cupressus L.). – Киев: Аграрна наука, 2006. –256 с.
Захаренко Г. С., Севастьянов В. Е., Салогуб Р. В. Репродуктивное развитие и качество семян плосковеточника восточного в Крыму // Труды Санкт-Петербургского НИИ лесного хозяйства. – 2022а. – № 1. – С. 34–46.
Захаренко Г. С., Севастьянов В. Е., Салогуб Р. В. Особенности репродуктивного развитие Рlatycladus orientalis (l.) Franco в Крыму // Бюллетень Главного ботанического сада РАН. – 2022б. – № 1. – С. 40–47.
Кузнецов С. И. Основы интродукции и культуры хвойных Древнего Средиземья на Украине и в других районах юга СССР. – Киев: Наукова думка, 1984. – 124 с.
Леопольд А. Рост и развитие растений. – М.: Мир, 1968, – 494 с.
Мамаев С. А. Формы внутривидовой изменчивости древесных растений (на примере семейства Pinaceae на Урале). – М.: Наука, 1973. – 284 с.
Некрасов В. И. Актуальные вопросы развития теории акклиматизации растений. – М.: Наука, 1980. – 101 с. Тахтаджян А. Л. Высшие растения. Т. 1. От псилофитовых до хвойных. – М.-Л., Изд. АН СССР, 1956. – 488 с. Яблоков А. В. Популяционная биология. – М.: Высшая школа, 1987. – 303 с.
Яблоков А. В., Ларина Н. И. Введение в фенетику популяций. Новый подход к изучению природных популяций. – М.: Высшая школа, 1985. – 159 с.
The Gymnosperm Database, http://www.conifers.org/cu/Platycladus.php (291.2017)
Guoqing L., Sheng D. and Zhongming W. Mapping the climatic suitable habitat of oriental arborvitae (Platycladus orientalis) for introduction and cultivation at a global scale // Scientific Reports. – 11. 2016. – Vol. 6. – P. 1–9.
Farjon, A., Filer D. An atlas of the World’s Conifers: An analysis of their distribution, biogeography, diversity and conservation status. – Leiden, Boston: Brill, 2013. – 524 p.
Fu L., Yu Y., and Robert R. M. Taxodiaceae. In Wu Zheng-yi and Peter H. Raven (eds.). Flora of China, Volume 4.
Beijing: 1999. Science Press; St. Louis: Missouri Botanical Garden. (http://www.efloras.org)
Hagerup О. Zur Organogenie und Phylogenie der Koniferen-Zapfen // Kongelige Danske Videnskabernes Selskab.
Biologiske meddelelser. – 1933. – B. 10. – S. 1–82.
Krüssmann G. Manual of Cultivated Conifers. – Tumber Press: Portland, Oregon, 1985. – 361 p.
Lemoine-Sebastian C. La Vaskularisation du complexe bractée-ecaille chez les Taxodiacées // Travaux du Laboratoire Forestier de Toulouse. – 1968. – Vol. 7, N 1. – P. 1–22.
Liu F., Xing S., Ma H., Du Z., Ma B. Cytokinin-producing, plant growth-promoting rhizobacteria that confer resistance to drought stress in Platycladus orientalis container seedlings // Applied microbiology and biotechnology. – 2013. – Vol. 97, N. 20. – 2013. – P. 9155–9164.
Jagel A., Dӧrken V. M. Morphology and morphogenesis of the seed cones of the Cupressacae – part II Cupressoideae // Bulletin of the Cupressus Conservation Project. – 2015, N 4 (2), – P. 51–78.
Sun Z., Qi T., Jin C., Wang Z. Famous ancient Trees in Beijihg, – Beijihg: Publishing House, 1992. – 126 s.
Yang X., Dong N., Yan D., Sajiki N., Zhao Y. The growth and distrubution of Platycladus orientalis seed-base seedling root in different culture periods // Shengtai xuebao. – Vol. 31, N 19. – 2011. – P. 5818–5823.
Wang L., Feng J., Wan X., Wang S., Jia C. The interaction of drought and slope aspect on growth of Quercus variabilis and Platycladus orientalis // Shengtai xuebao. – 2013. – Vol. 33, N 8. – P. 2425–2431.
Zakharenko G. S., Salogub R. V., Sevastyanov V. E. Variation of biomorphological characteristics of cones of oriental arborvitae (Platycladus orientalis (L.) Franco) in the Crimea // Ekosistemy. 2022. Iss. 31. P. 61–71.
The intraspecific variation of cones of oriental arborvitae (Platycladus orientalis (L.) Franco) has been studied in ornamental plantations in the Foothill Crimea and the South Coast of Crimea, as well as in four forest plantations in the coastal zone of Evpatoria. The endogenous, individual and interdemic variation of the structural features of cones has been determined: length and thickness in mutually perpendicular directions in the thickest part, the number of scales and seeds, the length of the apophysis of the most developed scales. At the individual level, the average statistical values of the parameters of the cones are: length – 13.8–22.6 mm (6 % ≤ C ≤ 17 %), larger diameter – 12.0–18.6 mm (7 % ≤ C ≤ 19 %), smaller diameter – 12.0–15.7 mm, number of scales – 5.9–7.6 pcs. (7 % ≤ C ≤ 14 %), apophysis length – 0.16–2.09 mm (29 % ≤ C ≤ 106 %), number of seeds – 4.5–7.3 pcs. (9 % ≤ C ≤ 36 %). The results obtained confirm the data on the organ specificity of the levels of variability of the structural elements of generative organs in gymnosperms. The wide range of variation in the length of the apophysis is associated with its atavistic nature due to the morphological appearance of the covering scale, fused with the seed scale at the early stages of evolution of modern species of the Cupressaceae family, and the conduplicated development of the scales of their cones. The levels of endogenous and individual variation coincide in oriental arborvitae, as in natural populations of Pinus sylvestris and Picea abies. This indicates a balanced phenetic polymorphism of the introduced Crimean population of Platycladus orientalis and indicates a high microevolutionary potential of the species in conditions of isolation and the possibility of formation in the southern regions of Russia during 20–25 overlapping seed generations of local reproduction of introduced populations comparable in terms of phenotypic variability to natural populations of gymnosperms.
Key words: Platycladus, variation, morphological features, cones, introduction.
Поступила в редакцию 17.10.22 Принята к печати 27.10.22
