
ИЗМЕНЕНИЯ РАЗМЕРНО-ВЕСОВЫХ ПОКАЗАТЕЛЕЙ ТЕЛА В ЗАВИСИМОСТИ ОТ СЕЗОНА И ПОЛА У НЕКОТОРЫХ РЫБ ЮГО-ВОСТОЧНОГО ПОБЕРЕЖЬЯ КРЫМА
CHANGES IN BODY SIZE-WEIGHT INDICES DEPENDING ON SEASON AND SEX IN SOME FISHES OF THE SOUTH-EASTERN COAST OF CRIMEA
JOURNAL: «EKOSISTEMY», Issue 39, 2024
Publication text (PDF): Download
UDK: 591.4:597.2/.5(262.5)
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Silkin Yu. A., Silkina E. N., Silkin M. Yu., Chernyaeva B. N.
T.I. Vyazemsky Karadag scientific station — Nature Reserve of the Russian Academy of Sciences, Feodosia, Russia
TYPE: Article
DOI: https://doi.org/10.29039/2413-1733-2024-39-95-104
PAGES: from 95 to 104
STATUS: Published
LANGUAGE: Russian
KEYWORDS: fish, size, weight, sex, season, Black Sea, South-Eastern Crimea.
ABSTRACT (ENGLISH):
The changes in body length and weight in fish of different natural mobility, depending on the season and sex of the individuals, were studied. It was revealed that seasonal changes in the size and weight indicators of the fish bodies are characterized by an increase in their values in autumn in bottom and demersal sedentary species (East Atlantic peacock wrasse (Symphodus tinca), scorpionfish). The belonging of the studied sample of sedentary species to the same population allows conducting studies on changes in body size-weight indices of the S. tinca and scorpionfish during their annual life cycle accurately. Female scorpionfish and male S. tinca had the highest size-weight indices during the season. For schooling, active fish (horse mackerel, smarida, weasel), this method is not correct as it is impossible to research one population in the studied sample during their migration. The studied sedentary species had more pronounced changes in body weight indices by season and sex. Thus, the weight indicators of sedentary fish were 1.5 times higher than their changes depending on sex. The obtained size-weight results confirm two fundamentally different feeding strategies in the S. tinca and scorpionfish. The sedentary scorpionfish uses an ‘intensive’ strategy. The possibility of implementing this strategy in the scorpionfish is due to the fairly high density of their potential prey per unit area of the species’ habitat. In the event of a drop in prey density, this strategy may be significantly ineffective and could result in a substantial limitation of opportunities for this fish species. The more mobile, bottom-dwelling S. tinca has an ‘extensive’ strategy due to a high intensification of food consumption and a more active metabolism. The extensive feeding strategy characteristic of the S. tinca implies the exploitation of larger areas of the coastal bottom seabed and, consequently, more intensive feeding throughout the entire annual cycle of fish. However, this method of foraging food by S. tinca requires high energy costs, which significantly affects the rate and magnitude of the increase in their body size and weight characteristics.
ВВЕДЕНИЕ
Методы морфофизиологических исследований в ихтиологии обычно используются для первичной, самой приблизительной оценки разных районов ареала обитания рыб. При сравнении таких показателей между собой можно определить границы благополучных и не благополучных условий существования тех или иных видов рыб. Изучение изменений размерно-весовых показателей тела рыб в зависимости от разных факторов является важной задачей, позволяющей решать ряд вопросов экологической и продукционной биологии гидробионтов, что становится особенно актуальным при усиливающейся антропогенной нагрузке на экосистемы и в условиях глобальных изменений климата нашей планеты. Одним из самых важных биологических ритмов в жизни рыб является их годовой жизненный цикл, различные сезоны которого, оказывают огромное влияние на их соматические и размерно-весовые характеристики (Шульман, 1972).
Морфофизиологические изменения рыб в разные периоды годового жизненного цикла, в пределах исследуемой акватории, позволяют выявить основные факторы, оказывающие влияние на их биологию. Выявление таких факторов, прежде всего кормового базиса рыб, как фундаментальной основы их существования, дает возможность оценить его соответствие биологическим потребностям исследуемых видов в исследуемом районе. Это важно для мониторинга биоразнообразия региона и трендов его развития на отдаленную перспективу.
Максимальному и интенсивному освоению кормовой базы рыбами способствуют приспособления, которые выработались в процессе эволюции. Наиболее действенным средством, позволяющим максимально освоить обширную кормовую базу для рыб, является движение. Высокая функциональная активность рыб, обеспечивающая освоение больших территорий, позволяет конкурировать с особями, не имеющих этого приспособления. Кроме влияния естественной подвижности, размерно-весовые показатели рыб, также зависят от пола. Различия этих показателей от пола редко принимаются во внимание несмотря на различия двигательных реакций самцов и самок у некоторых видов в период нереста.
Из литературных источников известно, что чаще всего размерно-весовые характеристики тела прибрежных видов рыб применяются в качестве индикаторов загрязнений среды обитания (Кузьминова, 2006; Руднева и др., 2010; Руднева и др., 2011; Чеснокова, 2017). Кроме того, загрязнение прибрежных вод оказывает большое влияние на генеративную функцию рыб (Руднева и др., 2010). Такие влияния могут изменять сроки созревания, и даже перерождение половых продуктов, что отражается на их морфологических показателях.
Цель настоящей работы состояла в изучении половых и сезонных изменений размерно-весовых характеристик тела у 5 видов черноморских рыб различной естественной подвижности, обитающих в прибрежье Карадагского природного заповедника в Юго-восточного побережья Крыма.
МАТЕРИАЛЫ И МЕТОДЫ
Объектами исследования были взяты пять видов рыб прибрежной акватории Юго-Восточного побережья Крымского полуострова: зеленушка рулена – Crenilabrus tinca Linnaeus, 1758; смарида – Spicara flexuosa Rafinesque, 1810; ласкирь – Diplodus annularis Linnaeus, 1758; скорпена – Scorpaena porcus Linnaeus, 1758; ставрида – Trachurus mediterraneus ponticus Aleev, 1956. Рыб отлавливали в прибрежье Карадагского заповедника, где особи были в наименьшей степени подвержены антропогенному воздействию. Отлов рыб осуществляли с помощью удочки и придонных ловушек в течение всего годового цикла (скорпена, ставрида) или только зимой, летом и осенью (смарида), или весной, летом и осенью (зеленушка рулена, ласкирь). Некоторые исследуемые виды (зеленушка рулена, смарида) имели вторичные половые признаки по цвету или размеру тела. По уровню естественной подвижности исследуемые виды были разделены на: донных малоподвижных (скорпена), придонных малоподвижных (зеленушка рулена), умеренно подвижных (смарида, ласкирь) и быстроплавающих (ставрида). Возраст всех выловленных для анализа видов рыб был в пределах 3–5 лет.
После отлова случайной выборки рыб определяли пол каждой особи, общую длину тела (TL) в см, вес тела без внутренностей (TW) в граммах. Полученные результаты были обработаны статистически. Определяли среднюю и квадратичную ошибку (М±m) (Лакин, 1990). Различия изменений размерно-весовых показателей тела рыб от сезона и пола выражались в процентах.
РЕЗУЛЬТАТЫ
Полученные результаты величин размерно-весовых характеристик тела у самцов и самок рыб, выловленных в различные периоды годового цикла представлены в таблице 1.
Исходя из данных по размерно-весовым показателям рыб, видно (смотри таблицу), что среди всех исследованных видов наибольшую (до 30 см) длину тела имела зеленушка рулена. Длина скорпены лишь немногим уступала зеленушке (до 28,5 см). У остальных видов длина тела была несколько меньше: у ставриды – до 23,0 см, у ласкиря – до 22,0 см, у смариды – до 19,8 см. Самой крупной по массе тела из исследованных видов была скорпена (308,6 г). У зеленушки масса тела также была высокой (до 298 г), за ней следовали ласкирь (до 186 г) и ставрида (до 98,6 г). Самый низкий вес тела был у смариды, вес самой крупной особи не превышал 69,0 г. Таким образом, среди исследованных рыб малоподвижные виды имели наибольшую длину и вес тела в отличие от более активных видов.
Полученные результаты сезонных и половых изменений исследуемых показателей рыб разной подвижности были объединены по исследуемым параметрам и представлены на рисунках 1 и 2.
Таблица 1
Величины веса и размера тела некоторых черноморских рыб в зависимости от пола и сезона
Таблица 1 (продолжение)
| 1 | 2 | 3 | 4 | 5 | 6 | 7 |
| ЛаскирьDiplodus annularis | ♂ | Весна | 16,5–21,5 | 19,0±1,2 n=14 | 102,0–186,0 | 135,0±12,0n=10 |
| Лето | 13,5–17,6 | 15,5±1,1 n=12 | 44,0–110,0 | 62,5±2,7 n=11 | ||
| Осень | 12,5–14,3 | 12,8±1,0 n=18 | 23,5–47,6 | 35,7±0,6 n=10,0 | ||
| ♀ | Весна | 13,1–19,2 | 17,5±1,2 n=15 | 103,5–147,6 | 116,2±1,2 n=10 | |
| Лето | 15,0–22,0 | 17,2±1,0 n=15 | 75,5–145,0 | 108,9±7,6 n=13 | ||
| Осень | 13,8–21,0 | 15,8±1,2 n=14 | 40,2–50,8 | 47,2±3,6 n=12 | ||
| СтавридаTrachurus mediterraneus ponticus | ♂ | Зима | 17,0–23,0 | 20,3±1,7 n=16 | 54,1–98,6 | 54,8±3,6 n=15 |
| Весна | 16,9–23,1 | 16,0±0,1 n=60 | 20,2–54,0 | 33,5±1,9 n=60 | ||
| Лето | 17,2–20,8 | 16,0±0,9 n=55 | 14,0–7,6 | 30,6 ±1,8 n=63 | ||
| Осень | 14,3–16,2 | 15,3±1,1 n = 17 | 16,7–37,7 | 31,4±1,4 n=15 | ||
| ♀ | Зима | 17,0–21,0 | 18,8±1,2 n=15 | 33,5–73,7 | 58,3±2,7 n=12 | |
| Весна | 14,0–20,0 | 15,3±0,9 n=38 | 15,4–33,1 | 20,3±1,2 n=35 | ||
| Лето | 13,0–15,5 | 14,4±0,9 n=109 | 17,0–37,8 | 26,2±1,6 n=109 | ||
| Осень | 13,0–15,5 | 14,4±0,9 n=27 | 20,9–38,4 | 27,7±0,01 n=17 |
Примечание к таблице: n – число рыб в опыте; TL – размер рыбы (см); TW – вес тела рыбы.
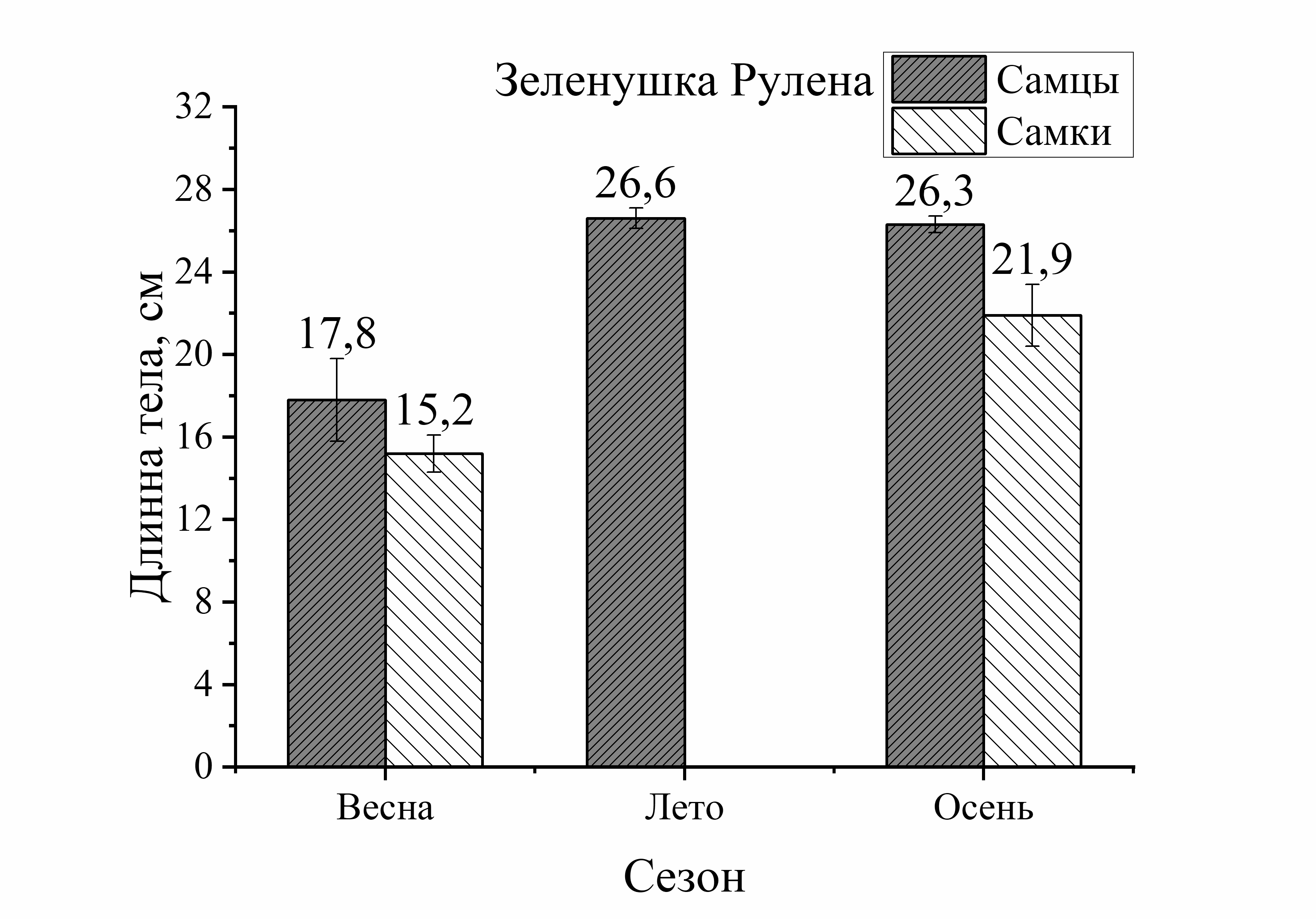
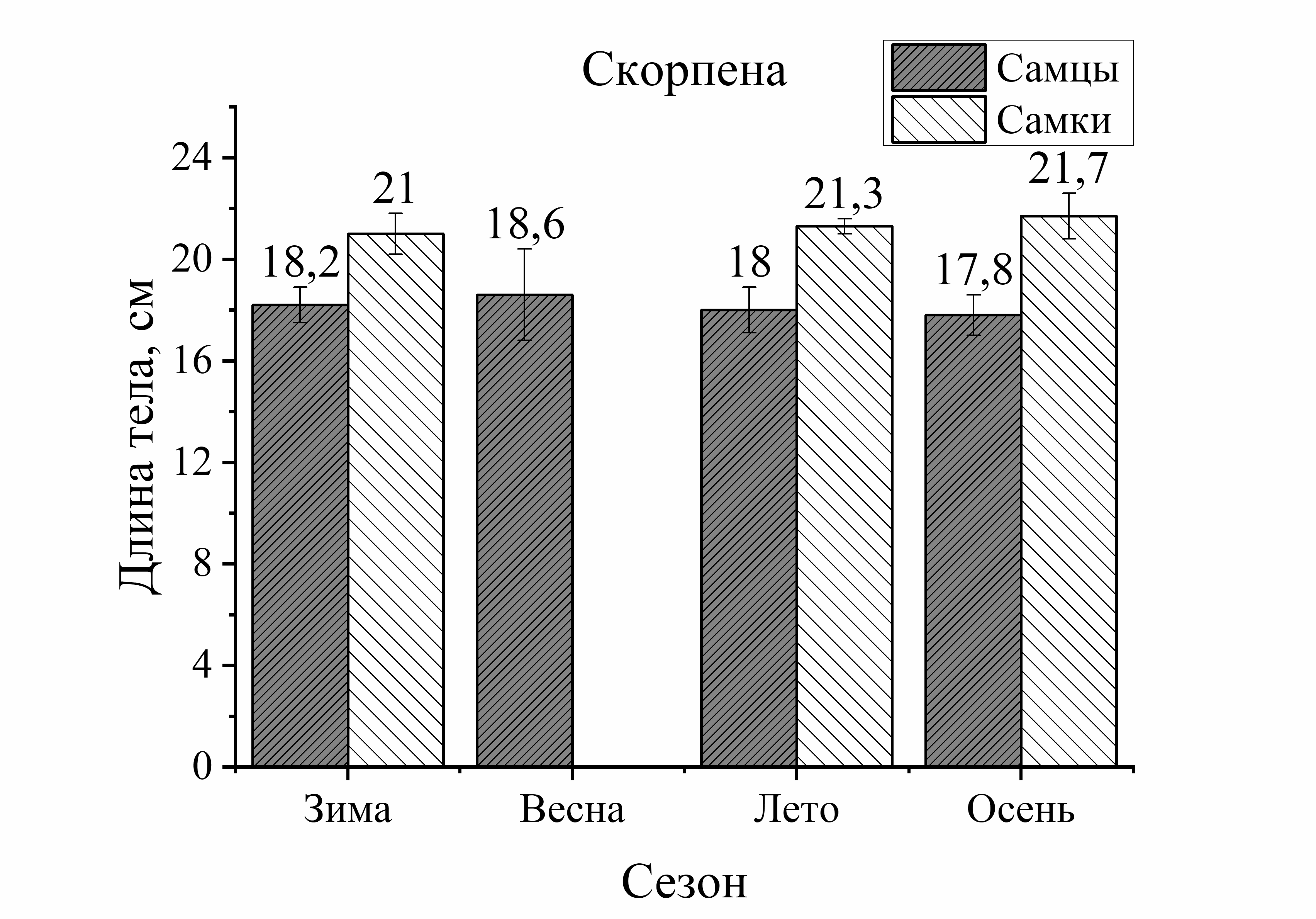
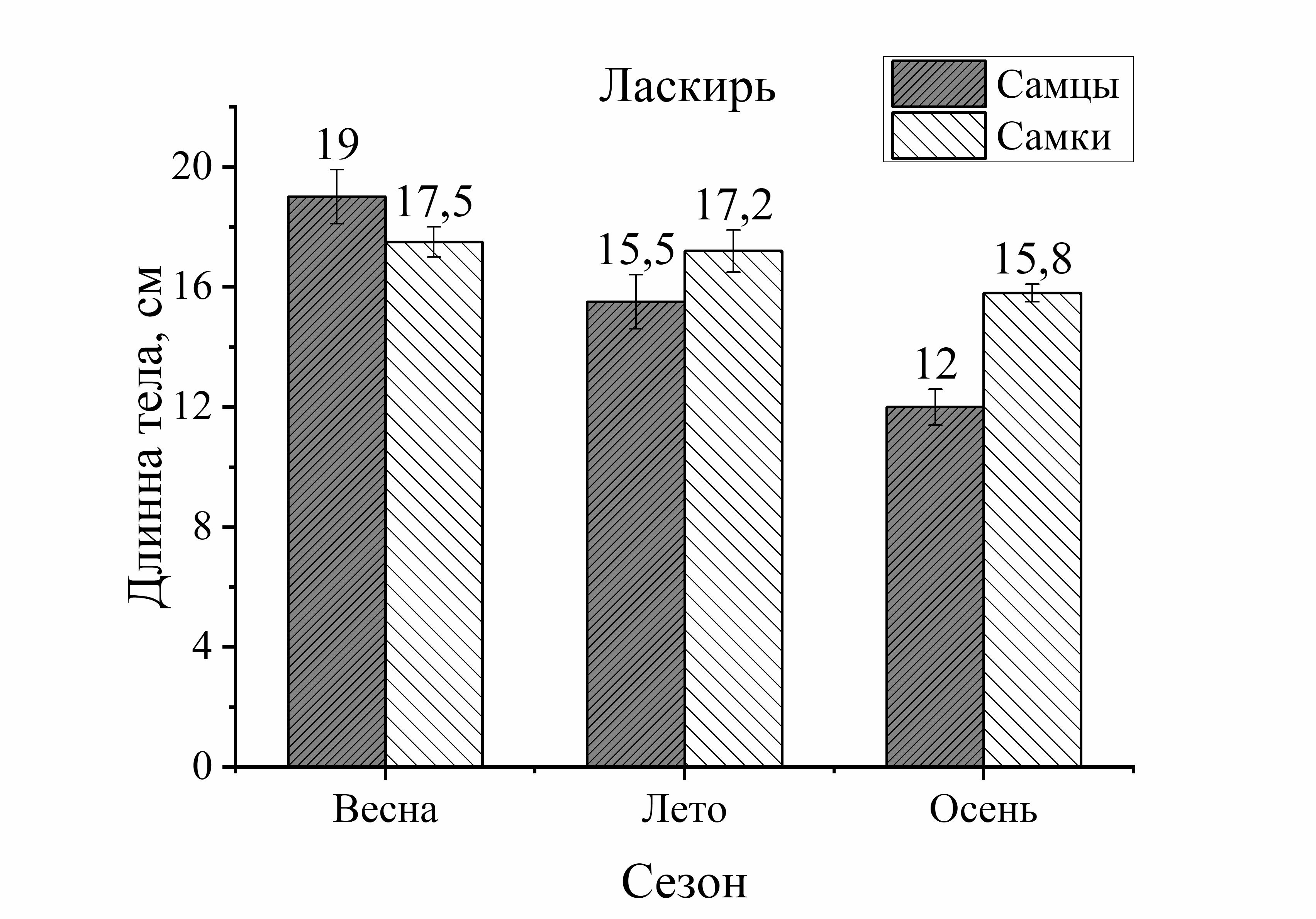
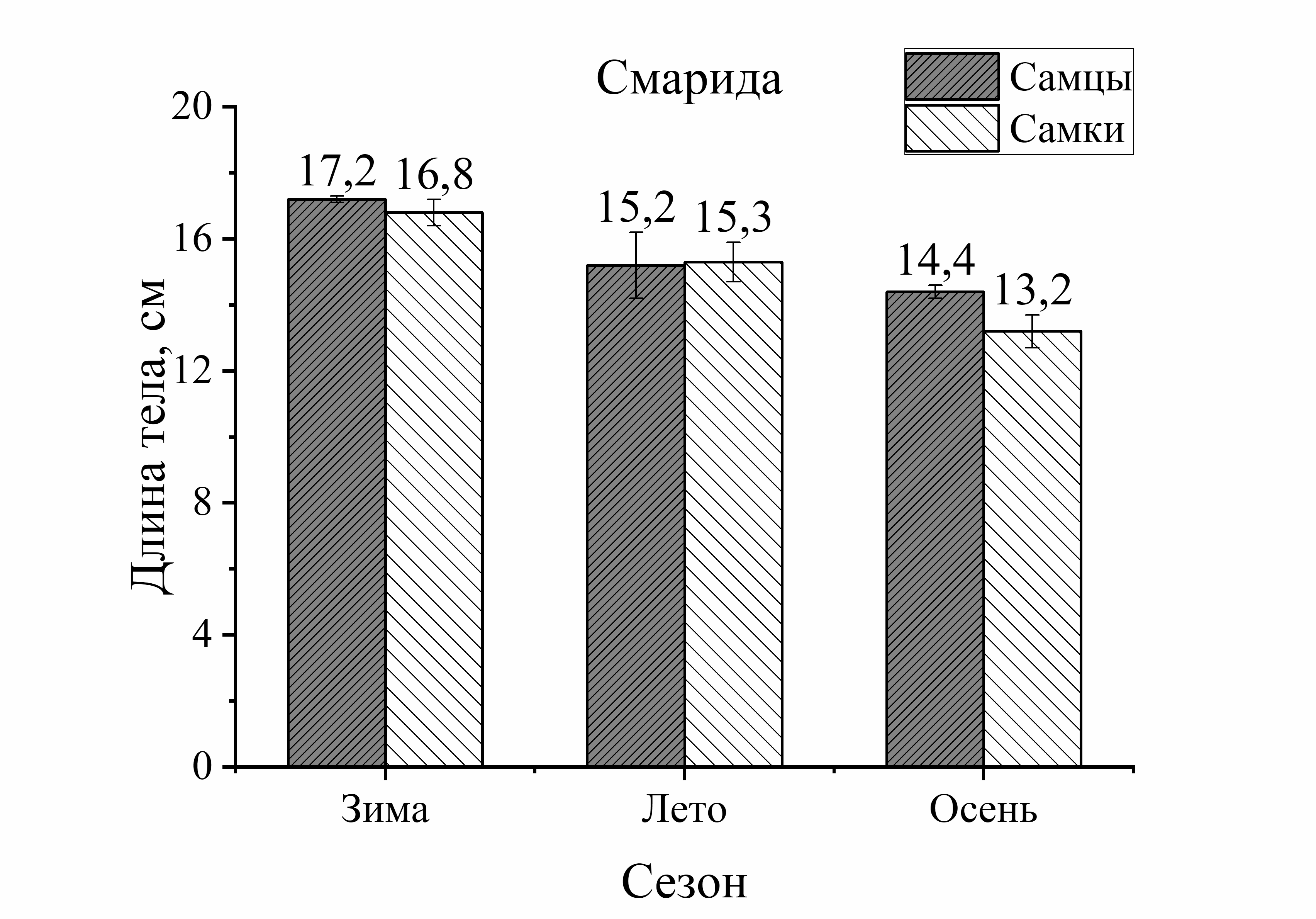
Из рисунка 1 видно, что сезонные изменения общей длины тела у выловленных активных видов рыб (смарида, ставрида) характеризовались понижением размера тела летом и осенью по сравнению с зимой. У ласкиря длина тела с весны до осени уменьшалась только осенью. Напротив, у зеленушки размер тела был выше летом и осенью по отношению к весне, а у малоподвижной скорпены этот параметр не имел достоверных изменений в течение годового цикла. Следует отметить, что наибольшие изменения длины тела у всех исследованных видов (кроме скорпены) были отмечены в осеннем периоде. Так, размер тела у зеленушки рулены осенью увеличивался у обоих полов на 32 % (по отношению к весне). У остальных видов рыб он уменьшался следующим образом: у ласкиря размер тела осенью был на 37 % у самцов и у самок – на 11 % меньше, чем весной. У смариды этот показатель был на 36 % у самцов и у самок на – 9 % ниже осенью, чем в зимний период. Длина тела ставриды у обоих полов была на 23–25 % меньше осенью, чем зимой. Таким образом, наибольшее увеличение длины тела особей, отловленных осенью было отмечено у зеленушки. Длина тела скорпены не изменялась от зимы к осени, а у ласкиря, смариды и ставриды этот показатель, в среднем, примерно одинаково уменьшился в осеннем периоде (рис. 1).
Изменения длины тела в зависимости от пола у исследованных видов были неоднозначны. Они выражались в преобладании размера тела самцов (на 15–17 %) у придонного вида – зеленушки рулены (лето – осень) и увеличении размера тела самок
 |
 |
 |
 |
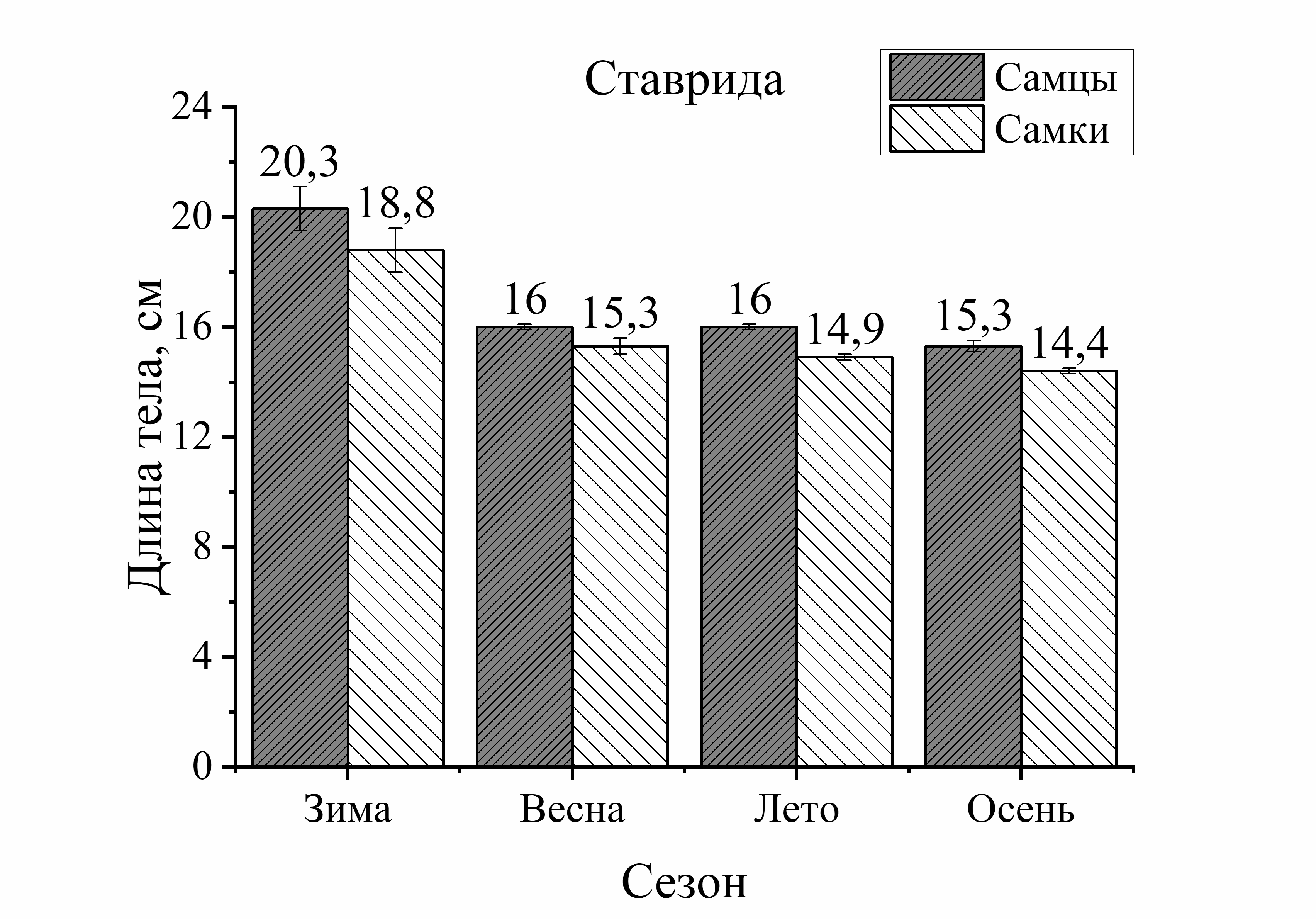 |
Рис. 1. Сезонные изменения общей длины тела в зависимости от пола рыб различной естественной подвижности
малоподвижного вида – скорпены (зима – осень). У скорпены размер тела самок в течение сезона был всегда выше длины самцов и в сравнении с другими видами эти изменения были наиболее выражены. Так, увеличение длины тела самок скорпены зимой составляло 13 %, летом – 23 %, а осенью – 18 %. У смариды в период зима – лето достоверных различий этого показателя от пола не было отмечено, лишь осенью длина тела самцов была на 19 % больше, чем у самок. Величина длины тела ставриды в течение года в зависимости от пола достоверно не изменялась. У ласкиря размер тела самок был на 14 % больше длины самцов летом, а осенью – на 24 %. Таким образом, в течение сезона преобладание общей длины тела самок над самцами было отмечено у скорпены и ласкиря, а самцов над самками – у зеленушки рулены и смариды. Наибольшие колебания значений длины тела от половой принадлежности были отмечены у малоподвижных и придонных видов, в отличие от умерено – и быстроплавающих видов (смарида, ставрида) (рис. 1).
Анализ сезонных и половых изменений длины тела был неоднозначным у исследованных рыб. Так, у зеленушки колебания (min–max) длины тела в течение сезона составили 16,0 см, а изменения длины от пола составили 13,0 см у самцов и 15 см у самок. В среднем изменения длины от пола составили – 14,0 см. То есть, сезонные изменения длины тела у зеленушки были в 1,1 раз больше изменений от пола. У скорпены подобные сопоставления показали, что сезонные изменения размера тела составили 14,5 см, а половые – 9,5 см у самцов и 13,0 см у самок (в среднем 11,5 см). То есть, сезонные изменения длины тела у скорпены были в 1,3 раза больше изменений от пола у этих особей. У ласкиря подобные расчеты показали, что сезонные изменения длины тела у этого вида не отличались от межполовых (1,1). У смариды и ставриды соотношения изменений длины тела от сезона и пола также мало различались (1,2 и 1,1, соответственно). То есть, у активно плавающих видов изменения длины тела от сезона и пола слабо различались, а у малоподвижных изменения от сезона были на 30 % больше, чем от пола.
Изменения веса тела от сезона и пола у исследованных черноморских рыб представлены на рисунке 2. Из рисунка 2 видно, что изменения в весе тела от сезона и пола были более выражены по сравнению с изменениями размерных показателей тела рыб (см. рис. 1 и 2).
 |
 |
 |
 |
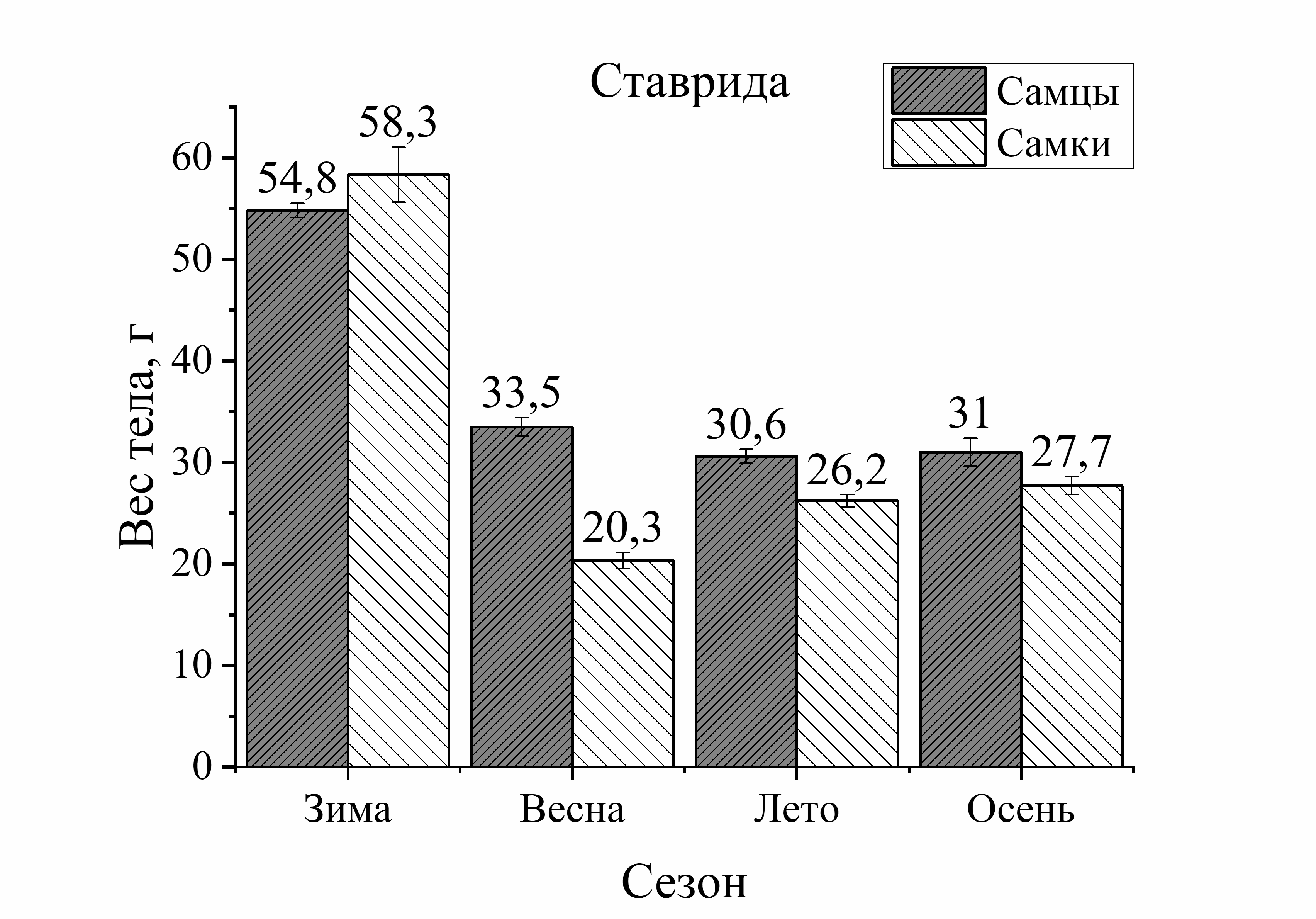 |
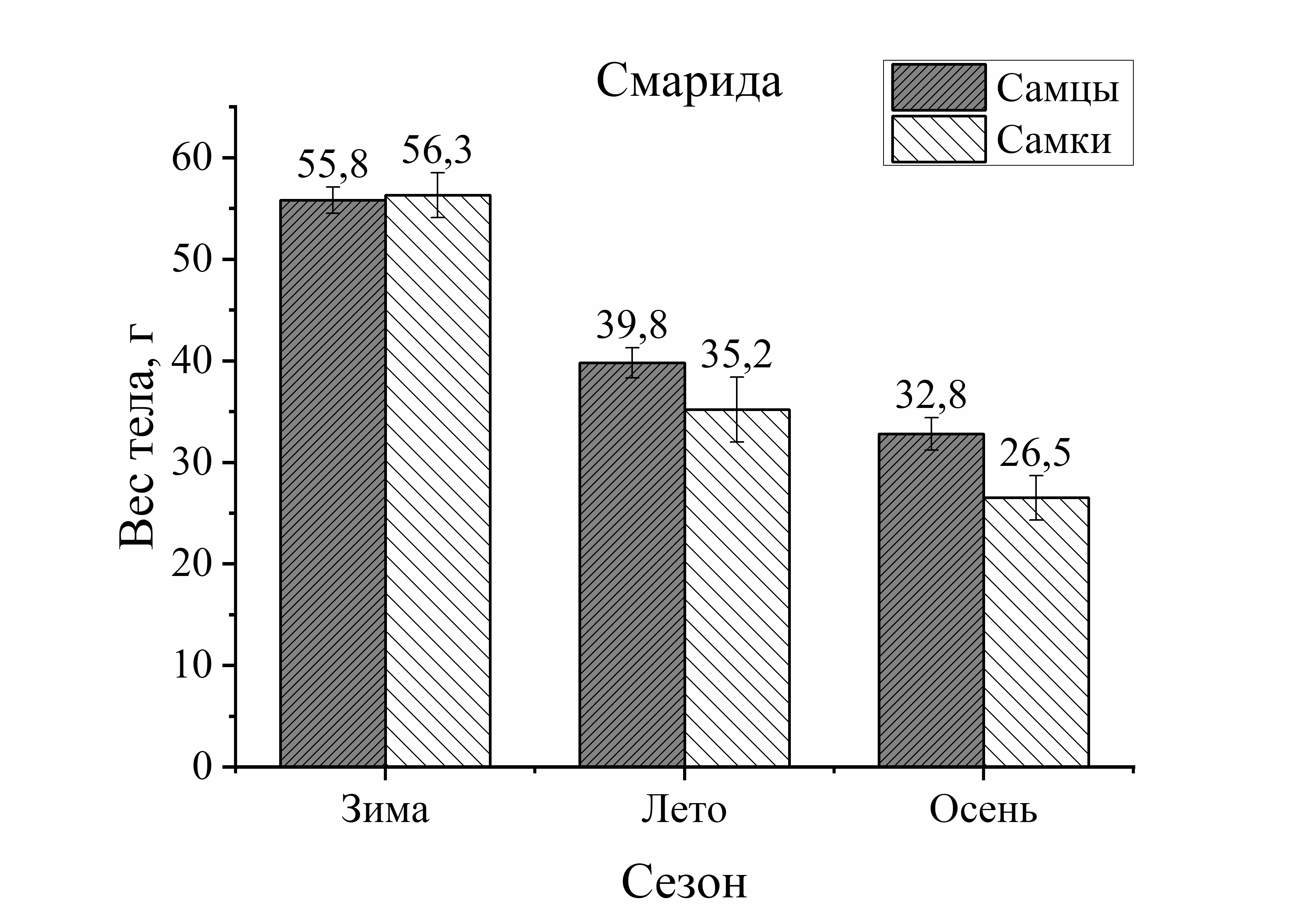
Рис. 2. Вес тела рыб различной естественной подвижности в зависимости от сезона и пола
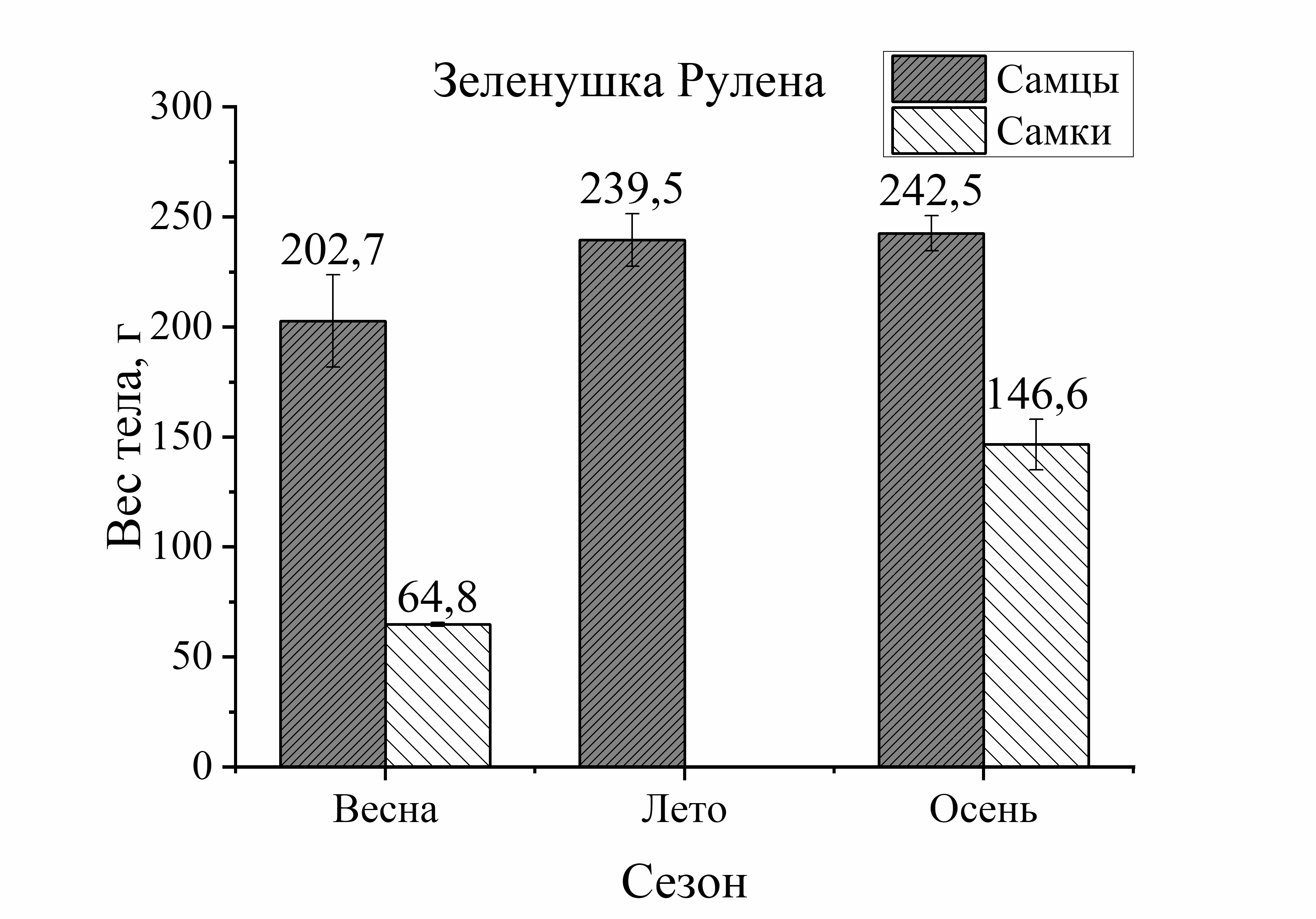
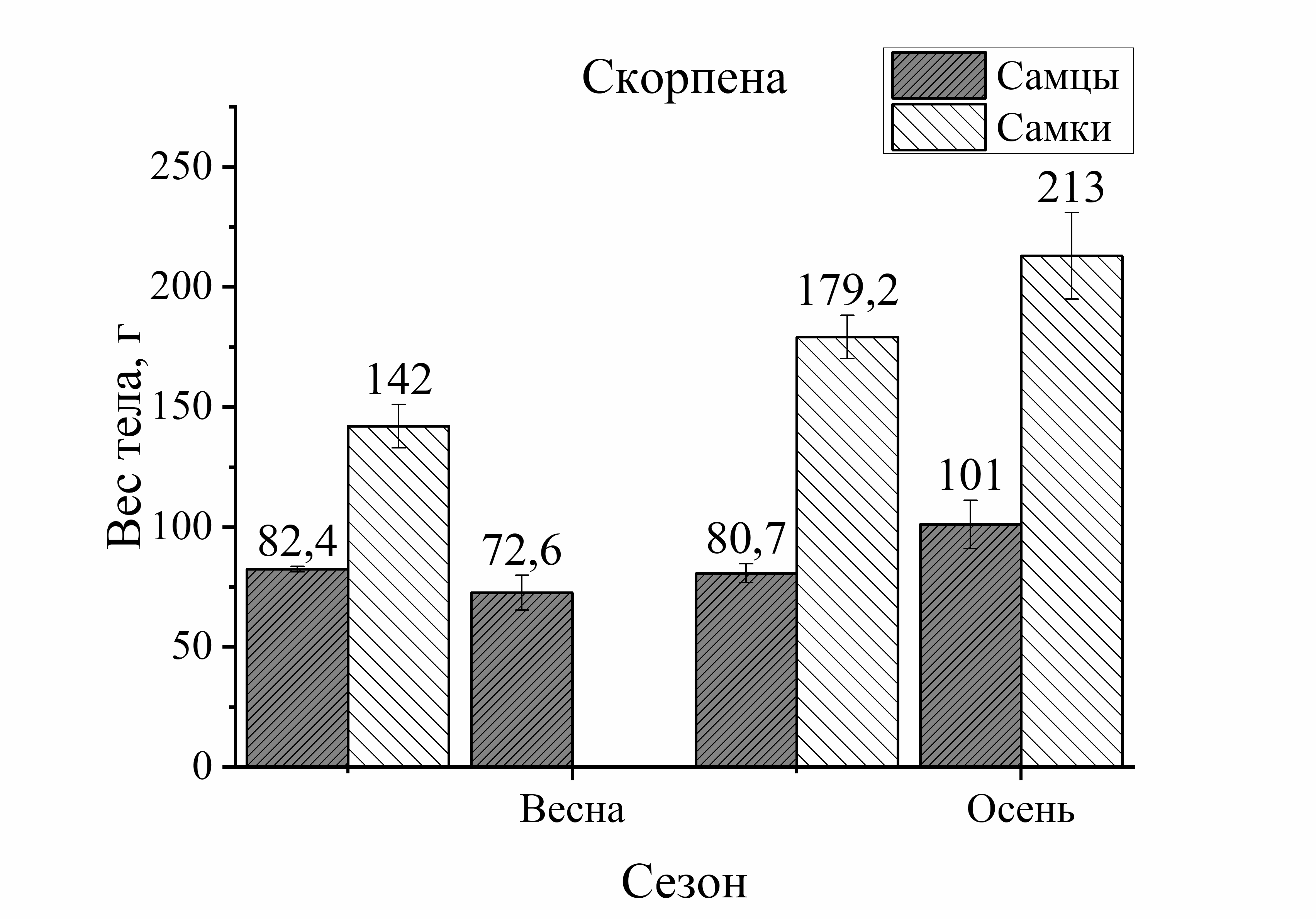
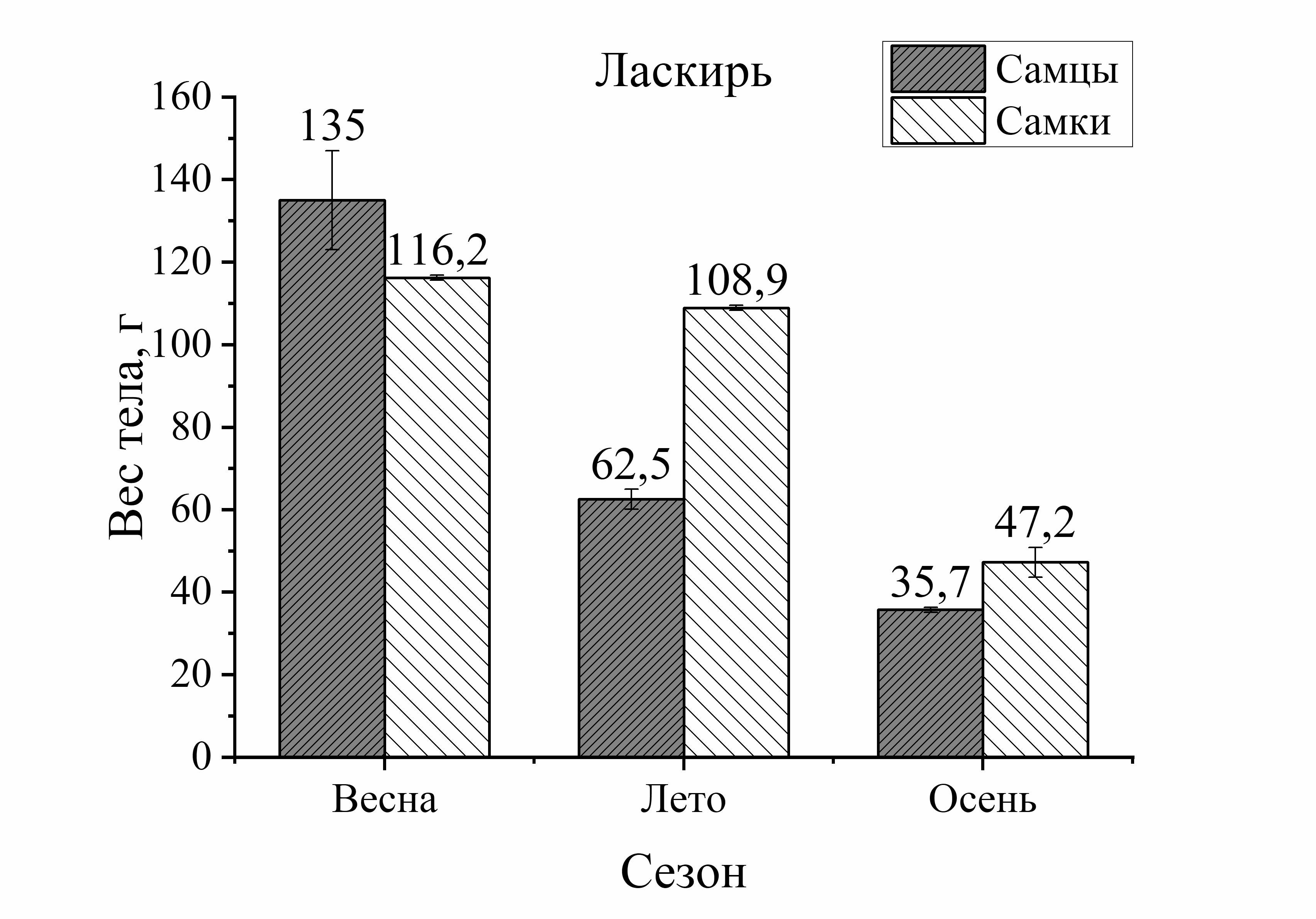
Так, вес тела у зеленушки рулены, отловленной осенью, был у самцов на 17 %, а у самок на 56 % больше по сравнению с весной (рис. 2). Вес тела скорпены с зимы к лету достоверно не изменялся у самцов, а у самок увеличивался на 21 %. Осенью этот показатель у самцов скорпены увеличивался на 18 %, у самок на 33 % по сравнению с зимними показателями. У других видов величина этой характеристики уменьшалась к осени. Так, вес тела у самцов ласкиря был ниже летом на 54 %, а осенью на 74 % по отношению к весне. У самок ласкиря вес тела летом достоверно не изменялся, а осенью снижался на 59 % по сравнению с весной (рис. 2). У смариды вес тела самцов уменьшался летом на 29 и на 15 % осенью, у самок этот показатель снижался более интенсивно: летом на 53 %, а осенью на 37 % по отношению к весне (рис. 2). Вес тела ставриды в период зима – осень, также как и у смариды, снижался более активно, чем у других видов. Так, вес тела у самцов ставриды был ниже на 39 % летом, осенью на 44 %, а у самок на 65 % летом и на 55 % осенью в отличие от зимы (рис. 2). Таким образом, сезонные изменения веса тела у рыб с меньшей двигательной активностью (зеленушка, скорпена) характеризовались увеличением веса тела осенью. Причем, наибольшее увеличение массы тела с весны к осени было отмечено у самцов зеленушки и у самок скорпены. У активно плавающих видов (смарида, ласкирь, ставрида) вес тела был значительно ниже осенью, по сравнению с зимне-весенним периодом. Причем, в большей степени снижение веса тела рыб в осеннем периоде было отмечено у самцов ласкиря и у самок ставриды и смариды.
Изменения веса тела от пола рыб были более интенсивно выражены у донных и придонных малоподвижных видов. Так, у зеленушки вес тела у самцов был на 68 % выше весной и на 40 % – осенью по сравнению с самками (рис. 2). У скорпены вес тела в течение всего сезона был выше у самок зимой – на 40 %, в летне-осеннем периоде – на 55 % по сравнению с самцами. У более активных рыб вес тела от пола изменялся в меньшей степени. Так, у ласкиря вес тела самцов был несколько выше веса самок только весной (на 14 %), а летом и осенью вес самок преобладал над таковым у самцов (на 43 и 21 %, соответственно) (рис. 2). У смариды вес тела от пола не изменялся зимой, а летом вес тела у самцов был на 12 %, а осенью на 44 % выше, чем у самок. У ставриды вес тела от пола изменялся аналогично изменениям у смариды. Так, в зимнем периоде достоверных изменений веса тела у ставриды от пола не было отмечено, а в остальные периоды года вес тела самцов ставриды был выше веса самок: весной – на 39 %, летом – на 14 %, осенью – на 11 % (рис. 2). Таким образом, зависимость веса тела рыб от пола в течение сезона характеризовалась преобладанием веса тела самцов над самками у зеленушки, смариды, ставриды и частично (весна) – у ласкиря.
Анализ изменений веса тела рыб от сезона и пола (смотри расчет взаимоотношений по длине тела) показал, что сезонные изменения веса тела были выше изменений от пола в 1,3 раза у зеленушки, в 1,7 раз – у скорпены, в 1,2 раза – у ласкиря, смариды и ставриды. Как видно, соотношения изменений по весу тела совпадают с изменениями по размерам тела. То есть, сезонные изменения размерно-весовых показателей у мало активных видов рыб значительно преобладают над изменениями от пола, в то время как у активно плавающих видов они были равнозначными.
ОБСУЖДЕНИЕ
При объяснении полученных результатов надо иметь в виду, что в опыт брались рыбы, отловленные в прибрежье, из которых оседлый образ жизни ведут только скорпена и зеленушка рулена. Это позволяет отнести эти виды к одной популяционной выборке, приуроченной к месту их вылова. Ставрида, смарида и ласкирь относятся к стайным видам рыб, и их отлов приурочен к периодам нереста и нагула в процессе миграционных перемещений вдоль побережья. В связи с миграционными перемещениями в течение года вдоль побережья отловленные виды из таких стай, скорее всего не относятся к одной популяционной выборке, а представляют разные популяции. С другой стороны, (смотри раздел «Материал и методы») выловленные для исследования рыбы были близки по возрасту, который у всех видов был в диапазоне 3–5 лет. Это было обусловлено способом вылова этих рыб: снасти, которые использовались для этих целей позволяли ловить рыб определенного размера и возраста. В связи с этим для исследованных видов, выловленных с одинаковым возрастным диапазоном, можно проводить сравнительные исследования ввиду их возрастной унификации. Для каждого оседлого вида рыб, выловленного из одной популяции, можно определить, относительно корректную зависимость исследуемых показателей от пола, уровня их двигательной активности и сезона. Для мигрирующих стайных рыб такой метод не является корректным ввиду невозможности работы с одной популяцией в выловленной выборке во время прибрежных миграций, о чем и свидетельствуют полученные результаты.
Изменения размерно-весовых показателей тела и органов рыб тесно связаны с физиологией организма и в основном отражают кормовую базу, выраженную степенью упитанности и темпами линейного прироста. Исследуемые виды рыб, в основном – зоофаги (зеленушка, смарида), а также истинные хищники (скорпена, ставрида). Объектами питания исследуемых рыб чаще всего являются другие, более мелкие виды рыб, а иногда – ракообразные. Исключением является ласкирь, основной пищей которого являются, водоросли, ракообразные, медузы, иногда – мелкая рыба (Болтачев, Карпова, 2017). В изменении длины и веса тела имеет значение частота приема пищи и процент ее усвояемости.
У скорпены в исследуемом периоде годового цикла (весна – осень) в уловах попадались рыбы одного размера, однако вес их особей был на 30 % больше осенью, чем весной (рис. 1, 2). Для скорпены пищей являются другие виды рыб и ракообразные, которые являются более калорийным кормом, чем пища у зеленушки. Известно, что скорпена, несмотря на то, что является хищником, в период нереста прекращает питание, даже может выдерживать перерывы в питании до 10–15 дней. Несмотря на неравномерность приема пищи, основной пик питания (жор) у скорпены приходится на лето и осень (Арнольди, Фортунатова, 1941; Васильева, 2007). В летне-осеннем периоде у прибрежных рыб, вследствие возрастания температуры среды, увеличивается интенсивность пищеварения за счет активизации пищеварительных ферментов. Это способствует увеличению интенсивности усвоения пищи у прибрежных рыб. Г. Шульман (1972) отмечал высокую сопряженность белкового роста в теле рыб, имеющих высокую обеспеченность пищей и малую продолжительность нереста.
Другой оседлый вид зеленушка, напротив, с весны до глубокой осени питается постоянно, причем неподвижным кормом (моллюски). Подход к берегам прибрежья более крупных по размеру и весу особей зеленушки и скорпены летом и осенью по отношению к весне видимо, обусловлен процессами интенсивного кормления и особенностями их нереста. В исследуемом периоде годового цикла (весна – осень) размер выловленных осенью особей зеленушки был на 30% больше, чем весной. При этом общий вес тела особей из случайной выборки, выловленных осенью самцов, был на 16 %, а у самок – на 50 % больше (табл. 1, рис. 1, 2), чем у рыб, отловленных в весеннем периоде. Отлов более крупных по размеру и весу особей зеленушки осенью, по отношению к весне видимо, и обусловлен особенностями процесса питания. Зеленушки осуществляют непрерывное питание постоянно присутствующим кормом – мелкими моллюсками, которые населяют заросли цистозиры и другие водоросли, что позволяет им не только увеличивать массу и дину тела, но и осуществлять полноценный гаметогенез. Этим и объясняется более высокие размерно-весовые характеристики, как у самцов, так и у самок в осеннем периоде по сравнению с весенними показателями. Образ жизни исследуемых придонных видов характеризуется не высокой естественной подвижностью при кормлении и осуществлении процесса нереста. Так, основным кормом для зеленушки является не очень высококалорийный корм (моллюски), но доступность к корму у этого вида постоянная, а скорость усвоения пищи у зеленушки в три раза выше, чем у скорпены (Арнольди, Фортунатова, 1941). Кроме того, плодовитость зеленушки рулены довольно скромная по сравнению с другими исследуемыми видами (от 12,4 до 57,8 тыс. икринок) (Васильева, 2007). Возможно, уменьшение энергетических затрат организма на движение и на обеспечение процессов продолжения рода, наряду с доступностью корма, дает основание к увеличению длины тела зеленушки рулены в период весна – осень.
Таким образом, малоподвижные рыбы (зеленушка рулена и скорпена) имеют положительную динамику прироста размерно-весовых характеристик в летне-осеннем периоде, что убедительно демонстрируют наши наблюдения (смотри табл. 1 и рис. 1, 2).
Мигрирующие виды рыб (ставрида, ласкирь, смарида), как показали наши исследования, не демонстрировали тренда в увеличении размерных и весовых характеристик тела в период весенне-летне-осеннего периода (рис.1, 2). У мигрирующих видов самые крупные по длине рыбы были выловлены у ставриды зимой: у ♂ L=17,0 – 23,0, у ♀ L=17,0–21,0) У ласкиря и смариды больший размер особей был в уловах весной (ласкирь с длиной тела у ♂ L=16,5–21,5, у ♀ L=12,5–19,7) или летом (смарида с размером тела у ♂ L=14,0–16,0 и у ♀ L=12,0–18,0). Осенью преобладали особи меньшего размера у подвижных видов: у ставриды самцы и самки были на 25 % меньше по сравнению с зимой, у ласкиря самцы были на 37 % и на 10 % – самки, и на 14 % размер самок смариды был меньше, чем весной (см. табл. 1). Такой результат не соответствует особенностям нагула у активно плавающих видов рыб. Так, ставрида является постоянно питающимся хищником в течение года и не прекращает питание даже в период нереста. Это быстро плавающая рыба, обладая высоким уровнем обмена, находится в постоянном движении при миграции, охоте, и бегстве от хищников (Беляев и др., 1983; Болтачев, Карпова, 2017). Не менее активными являются и другие мигранты. Ласкирь и смарида, как свидетельствуют ихтиологические наблюдения, также отличаются высокой интенсивностью питания (жор), приходящейся на осенние месяцы (Световидов, 1964; Васильева, 2007), в период которых необходимо было бы наблюдать повышение их размерно-весовых характеристик тела. Однако, нами такие результаты получены не были. Согласно нашим данным, вес тела активно плавающих видов в осеннем периоде характеризуется значительным снижением (на 53–59 %), чаще всего это отмечено у самок. У ласкиря отмечено даже не только резкое уменьшение веса у самок, но и у самцов (на 74 %). Такие результаты легко объяснить, если учесть присутствие в миграционных стаях разных популяций у активно плавающих видов. Так, по наблюдениям рыбаков было отмечено, что летом к берегу подходит мелкий ласкирь, в конце августа ловились крупные особи, а в сентябре – опять мелкий ласкирь (Световидов, 1964). Эти же наблюдения относятся к ставриде и смариде. Поэтому размерно-весовые показатели тела мигрирующих особей из случайной выборки, состоящей из смешанных сообществ и, естественно, имеющих разный вес и размер тела, не могут быть характеристикой тренда морфологических показателей этих видов в том виде, который вполне корректен для малоподвижных, оседлых видов рыб. Для этих видов требуется другой способ оценки, который опирался бы на выборку рыб из одной популяции (стаи) в разные периоды ее годового жизненного цикла. Разработка современных методов маркирования рыб могли бы существенно помочь в решении таких задач.
ЗАКЛЮЧЕНИЕ
Вылов одновозрастных оседлых и мигрирующих рыб в строго определенном месте и использование морфометрических методов оценки размерно-весовых трендов в разные периоды их годового жизненного цикла свидетельствуют о том, что эти методы с учетом миграционного фактора рыб имеют ограниченную область применения. Для зеленушки рулены и скорпены это вполне подходящий способ определения за некий временной интервал темпов прироста их размерно-весовых характеристик и интегральной оценки благополучия условий региона их оседлого проживания. Для мигрирующих видов (ставрида, смарида, ласкирь) такой способ оценки не применим ввиду невозможности получения для работы материала одной популяции в исследуемой выборке.
Сезонные изменения размерно-весовых характеристик оседлых рыб свидетельствуют о зависимости этих показателей не только от кормовой базы, но и от других факторов. К таким факторам в данном исследовании относились половая принадлежность и уровень двигательной активности. Так, самцы и самки зеленушки и скорпены по размерно-весовым характеристикам имеют «реципрокные» отношения. Более крупные самки скорпены порционно продуцируют значительно большее количество икры, упакованной в слизистые оболочки. Такая репродуктивная «активность» в значительной степени определяет более крупные размерно-весовые параметры самок скорпены по сравнению с их самцами. К тому же, как хищник-засадник скорпена питается в репродуктивный период активно, заглатывая рыбу и других гидробионтов, сопоставляемых, или даже больших, по ее размеру (Болтачев, Карпова, 2017).
У зеленушек напротив, много более крупные самцы «маскируют» достаточно невысокую репродуктивную активность. Мелкие моллюски – митилиды (Mytilidae, Rafinesque, 1815), которыми кормятся эти рыбы по своей питательной ценности менее продуктивны, но постоянное их употребление в пищу, видимо, и обеспечивает достаточно хорошие темпы прироста тела этих рыб за весенне-осенний период (см. табл. 1).
Полученные результаты отражают разную стратегию питания исследованных оседлых видов рыб (Шульман, Урденко, 1989). У хищника-засадника скорпены – это «интенсивный» путь питания. Возможность осуществления «интенсивной» стратегии для рыб хищников-засадников обусловлена достаточно высокой плотностью на единицу площади обитания этих видов, их потенциальных жертв. В случае падения этого показателя эта стратегия может существенным образом оказаться неэффективной и приводить к существенному ограничению возможностей для этих малоподвижных видов.
Экстенсивная стратегия питания, свойственная зеленушке, подразумевает питание рыб на более обширных пространствах прибрежного дна. Однако, такой способ добывания пищи требует высоких энергетических затрат, что существенным образом сказывается на темпах и величине прироста их размерно-весовых характеристик у рулены (Болтачев, Карпова, 2017).
Таким образом, эти две стратегии имеют свои слабые и сильные стороны, что и обеспечивает им право на существование, поскольку обе они способствуют биологическому прогрессу рыб.
Работа выполнена в рамках темы НИР «Изучение фундаментальных характеристик морских гидробионтов, обеспечивающих их функционирование в экосистемах и служащих основой их рационального использования и сохранения». Регистрационный номер 124030100100-0
- Арнольди Л. В., Фортунатова К. Р. К экспериментальному изучению питания рыб // Труды зоологического Института АНСССР. – 1941. – VII. – С. 44–93.
- Беляев В. И., Николаев В. М., Шульман Г. Е., Юнева Т. В. Тканевый обмен у рыб. – Киев: Наукова думка, 1983. – 144 с.
- Болтачев А. Р., Карпова Е. П. Морские рыбы Крымского полуострова. – Симферополь: Бизнес-Информ, 2017. – 376 с.
- Васильева Е. Д. Рыбы Черного моря. – М.: Изд-во ВНИРО, 2007. – 238 с.
- Кузьминова Н. С. Оценка токсического действия хозяйственно-бытовых сточных вод на морские организмы: автореф. дис. … канд. биол. наук: спец. 03.00.16 Экология. – М.: МГУ, 2006. – 23 с.
- Лакин Г. Ф. Биометрия. – М.: Высшая школа, 1990. – 352 с.
- Руднева И. И., Шевченко Н. Ф., Залевская И. Н. Сравнительный анализ некоторых биомаркеров самок и самцов морского ерша Scorpaena porcus Linne (Scorpaenidae), обитающего в условиях хронического антропогенного загрязнения // В сборнике научных статей «Современные проблемы физиологии биохимии водных организмов. Том I. Экология, физиология и биохимия водных организмов. Российская академия наук, Отделение биологических наук РАН, Программа фундаментальных исследований на 2009–2011 гг.: «Биологические ресурсы России: оценка состояния и фундаментальные основы мониторинга», Институт биологии КарНЦ РАН; Ред. кол.: Немова Н.Н., Чуйко Г.М., Мещерякова О.В., Мурзина С.А.— Петрозаводск КарНЦ РАН, 2010. С. 261–265.
- Руднева И. И., Скуратовская Е. Н., Дорохова И. И. и др. Биоиндикация экологического состояния морских акваторий с помощью биоиндикаторов рыб // Водные ресурсы. – 2011. – Т. 38, № 1. – С. 92–97.
- Световидов А. Н. Рыбы Черного моря. – М.–Л.: «Наука», 1964. – 551 с.
- Чеснокова И. И. Биомаркеры черноморских рыб как показатели экологического состояния среды их обитания: автореф. дис. … канд. биол. наук. 03.02.17 Гидробиология. – Севастополь, 2017. – 22 с.
- Шульман Г. Е. Физиолого-биохимические особенности годовых циклов рыб. – М.: Пищевая промышленность, 1972. – 368 с.
- Шульман Г. Е., Урденко С. Ю. Продуктивность рыб Черного моря. – Киев: Наук. Думка, 1989. – 188 с.
