
ГНЕЗДОВАНИЕ СМОЛЯНОЙ ПЧЕЛЫ MEGACHILE (CALLOMEGACHILE) SCULPTURALIS SMITH, 1853 (HYMENOPTERA, APOIDEA, MEGACHILIDAE) В УЛЬЯХ ФАБРА. ВЫБОР ПОЛОСТИ ДЛЯ ГНЕЗДА
NESTING OF THE RESIN BEE MEGACHILE (CALLOMEGACHILE) SCULPTURALIS SMITH, 1853 (HYMENOPTERA, APOIDEA, MEGACHILIDAE) IN FABRE HIVES. CHOISE OF A CAVITY FOR THE NEST
JOURNAL: «EKOSISTEMY», Issue 39, 2024
Publication text (PDF): Download
UDK: 595.799:591.563
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Ivanov S. P.1, Pivovarenko N. A.2
1V.I. Vernadsky Crimean Federal University, Simferopol, Russia
2Small Academy of Sciences of Crimea «Iskatel», Simferopol, Russia
TYPE: Article
DOI: https://doi.org/10.29039/2413-1733-2024-39-155-166
PAGES: from 155 to 166
STATUS: Published
LANGUAGE: Russian
KEYWORDS: Megachile sculpturalis, nesting, choice of а cavity for the nest, Fabre hive.
ABSTRACT (ENGLISH):
The article presents the results of an experimental study of the behavior of the resin bee Megachile sculpturalis Smith, 1853 inhabiting Fabre hives. The hives provided to the bees for colonization were made of segments of reed stalks (Phragmites australis) in the form of tubes, open on one side and closed on the other. The tubes were tied into bundles and inserted into the front wall of the hive or fixed to its sides. In addition, separate tubes were inserted into the front wall of the hive at different distances from each other. Female bees preferred to colonize tubes located on the front wall of the hive separately at a distance of 2.5 and 4.5 cm from each other (49.5 % and 40.7 % of colonized tubes, respectively). Tubes tied into bundles were colonized less willingly. Bundles located on the front wall of the hive were colonized more readily (32.0 % of the colonized tubes in bundles) than bundles located on the sides of the hive (12.8 % of the colonized tubes in bundles). Bunches assembled from tubes with a straight cut of the front edge were populated 2.3 times more often than bundles assembled from tubes with an oblique cut of the front edge. Tubes of the outermost row of a bundle of tubes were populated more often than tubes located in the second row or in the center of the bundle of tubes. The percentage of colonization of tubes in the first row was 16.5 %, in the second row – 13.5 %, and of tubes located in the center of the bundle – 7.7 %. The lower density of tube colonization in certain variants of the experiment is explained by the fact that in these variants the female bees had difficulties remembering the location of the tube they had chosen for colonization. The female bees populated tubes of different diameters in the range from 7 to 16 mm, without а preference for tubes of any diameter. The length of the populated tubes varied from 7 to 35 cm. А preference for tubes in any range of length was also not revealed. The lack of skills in assessing the linear parameters of the cavity for the nest in females of M. sculpturalis is explained by their loss due to the transition to a tenantry – а settlement of nests of other bee species. This assumption is confirmed by the known tendency of M. sculpturalis to populate holes of abandoned nests of Xylocopa bees, including in the Crimea. Tubes with a smooth concave bottom were populated 2.7 times more often than tubes with an uneven and convex bottom. This feature of nesting of M. sculpturalis is also interpreted as a consequence of the transition to a tenantry.
ВВЕДЕНИЕ
Гигантская смоляная пчела Megachile (Callomegachile) sculpturalis Smith, 1853 (Apoidea, Megachilidae) – коренной обитатель Юго-восточной Азии – в конце 20 века и начале 21 была случайно завезена в Северную Америку (Mangum, Brooks, 1997; Hinojosa-Díaz, 2008; Parys et al., 2015 и др.), а затем и в Европу (Quaranta et al., 2014; Lanner et al., 2021 и др.). В Крыму пчелы M. sculpturalis были обнаружены 2018 году (Ivanov, Fateryga, 2019; Ivanov et al., 2021).
Пчела M. sculpturalis – один из самых крупных видов пчел-мегахилид. Самки могут достигать длины тела до 27 мм. В Европе вид активен с июня по сентябрь, с региональными вариациями в зависимости от климатического режима.
В пределах первичного ареала вид считается полилектом (Kakutani et al., 1990; Maeta et al., 2008). В Европе M. sculpturalis отмечена на более чем 25 видах кормовых растений (Quaranta et al., 2014), однако идентификация пыльцы из ячеек гнезд показала присутствие в них только (или в основном) пыльцы софоры японской – Styphnolobium japonicum (L.) Schott (Westrich et al., 2015; Aguado et al., 2018; Andrieu-Ponel et al., 2018). По нашим наблюдениям в Крыму самки пчел M. sculpturalis используют для провиантирования ячеек только пыльцу цветков софоры (Ivanov et al., 2021).
Для устройства гнезд самки используют различные полости, которые они разделяют на отдельные ячейки перегородками из смолы деревьев и замазки из влажной земли (Maeta et al., 2008; Aguado et al., 2018; Le Féon, Geslin, 2018 и др.). Полости чаще всего представляют собой каналы, выгрызенные другими насекомыми в древесине (Bila Dubaić et al., 2021). Отмечены случаи активного вытеснения смоляной пчелой из своих гнезд пчел-плотников Xylocopa virginica (L.) (Roulston, Malfi, 2012; Laport, Minckley, 2012).
По нашим наблюдениям самки M. sculpturalis способны заселять полости из разных материалов и разных размеров в широком диапазоне. В данном сообщении мы ограничились представлением результатов изучения особенностей заселения пчелами M. sculpturalis отрезков стеблей тростника.
Цель наших исследований – изучить приоритеты самок M. sculpturalis при выборе полостей для строительства гнезд в отношении размеров полостей и их расположения относительно друг друга на передней стенке улья Фабра.
МАТЕРИАЛ И МЕТОДЫ
Для достижения поставленной цели самкам пчел M. sculpturalis был предложен для заселения улей Фабра. Устройство и ход изготовления улья отражены на рисунке 1 a–d.
В качестве полостей для строительства гнезд в ульях пчелам предлагали отрезки тростниковых стеблей (Phragmites australis (Cav.) Trin. ex Steud.) в виде трубок, открытых с одной стороны и закрытых с другой. Трубки вставляли в отверстия, просверленные в передней стенке улья (рис. 1 а, b). Одну треть трубок (38 трубок) вставляли в отверстия, просверленные в передней стенке улья на расстоянии 4,5 см друг от друга, еще треть (38 трубок) – вставляли в отверстия, просверленные на расстоянии 2,5 см друг от друга (указано расстояние между центрами отверстий). Еще 38 трубок были связаны в два пучка, которые были вставлены в два больших отверстия в передней стенке улья (рис. 1 a, b, d). Кроме того, еще 25 пучков трубок были закреплены по бокам передней стенки и под крышей улья на его «чердаке» (рис. 1d). Трубки в пучках располагались вплотную друг к другу. В этом случае среднее расстоянии между центрами входных отверстий трубок составляло 1,0 см.
Подсчеты заселенных трубок проводили после окончания периода гнездования. Отдельно считали трубки, расположенные на передней стенке улья на разном расстоянии друг от друга и отдельно в пучках, вставленных в стенку улья и расположенных по бокам улья.
В нашем эксперименте пчелам предлагали для заселения отрезки стеблей тростника двух видов – с прямым срезом переднего края и с косым срезом, при этом каждый из пучков составляли из трубок либо только с косым, либо только с прямым срезом (рис. 2b, 3 b, с). Подсчеты заселенных трубок в пучках, составленных из трубок с косым и прямым срезом, проводились так же, то есть отдельно.
Каждый из пучков трубок (за небольшими исключениями) составляли из 19 трубок. Это количество трубок обеспечивало идеально плотное прилегание трубок друг к другу, особенно при небольших отличиях в диаметре (рис. 4). При такой упаковке внешний ряд трубок пучка содержал 12 трубок, второй ряд – 6, и 1 трубка занимала центральное положение в пучке. Такое расположение трубок было использовано для оценки способности пчел к ориентации, то есть способности запоминать расположение выбранной для заселения трубки среди других трубок в пучке.
Суть этой методики состоит в следующем. Если предположить, что местоположение трубки из крайнего ряда запомнить легче, чем трубки из второго ряда и, тем более, центральной трубки, то по частоте заселения трубок разных рядов пучка можно судить о способности пчел того или иного вида запоминать расположение заселенной трубки. Равномерное заселение рядов свидетельствует о достаточно хорошей способности пчел запоминать расположение гнезда, а преимущественное заселение крайнего внешнего ряда – плохой.

Рис. 1. Изготовление экспериментального улья Фабра (а–d)
и самки Megachile sculpturalis, заселившие улей (e, f)
Хорошо видны отдельные трубки и пучки трубок, вставленные в переднюю стенку улья (а, b), корпус улья с приоткрытой задней дверцей (c) и улей, установленный на месте постоянных наблюдений за гнездованием пчел (d). Самка, заселившая трубку с косым срезом, вставленную в переднюю стенку улья (е), и самка, заселившая трубку с косым срезом в составе пучка (f), ниже видны трубки с прямым срезом в составе другого пучка.
Эти рассуждения основывается на наблюдениях за поведением самок при поиске трубки для заселения. После посещения нескольких трубок и выбора наиболее подходящей, самка не сразу приступает к доставке строительного материала. Она несколько раз отлетает от улья и возвращается в выбранную трубку, запоминая ее расположение. Если самке не удается несколько раз безошибочно обнаружить выбранную трубку, она прекращает ее поиски и начинает поиски другой трубки.
При разрезании стебля тростника на отдельные отрезки срез можно сделать над или под узлом, разделяющим междоузлия. В первом случае дно внутренней полости стебля будет полукруглым, вогнутым и гладким – верхняя полутрубка на рисунке 2c. Во втором случае дно

Рис. 2. Отрезки стеблей тростника, подготовленные
для помещения в улей и строение внутренней полости стебля тростника
Вид пучков, составленных из трубок с косым и прямым срезом, анфас (а) и под углом в 45° (b); вскрытые (c) и невскрытые (d) трубки, одна из которых (верхняя) была получена после среза стебля над узлом, разделяющим междоузлия, а вторая (нижняя) – под узлом. Крупный план узлов, вскрытых (e) и невскрытых (f) отрезков стебля, срезанных над (левый отрезок) и под (правый отрезок) узлом, разделяющим междоузлия.
будет не гладким и выпуклым – нижняя полутрубка на том же рисунке. В наших экспериментах число трубок, полученных разрезанием стебля над или под узлом, разделяющим междоузлия, было примерно одинаково. Заселенные трубки с ровным и неровным дном подсчитывали отдельно. Предварительно обращаем внимание на то, что такое вогнутое дно имеет форму идентичную форме дна каналов, выгрызаемых пчелами-плотниками в стволах деревьев (рис. 5).
Пчелам предлагали для заселения отрезки стеблей длинной от 5 до 35 см и диаметром от 7 до 16 мм (указана величина наружного диаметра). Стебли тростника, как было отмечено выше, нарезали на отрезки двумя способами – либо срезом стебля под узлом, разделяющим

Рис. 3. Гнездование Megachile sculpturalis в ульях Фабра
a – самка входит в гнездо, устроенное в трубке с косым срезом; b и с – пучки из трубок с косым и прямым срезом.

Рис. 4. Заселение пчелами Megachile sculpturalis трубок в пучках
а – вид пучка трубок, закрепленного на передней стенке улья и составленного из трубок с прямым срезом, видны 6 запечатанных гнезд в трубках под номерами 76, 111, 116, 117 и 122; b – схема взаимного расположения трубок в идеальном пучке, выделены трубки внешнего ряда (сетчатая штриховка), второго ряда (заливка серым цветом) и трубка в центре пучка (без выделения); с – результаты заселения трубок первого ряда, второго ряда и центральной трубки в пучках экспериментального улья.
междоузлие, либо – над узлом. И в том и другом случае длина отрезков стеблей определялась длиной междоузлий. В результате распределение отрезков, как по длине, так и по диаметру соответствовало нормальному распределению (распределению Гаусса), то есть с большей представленностью отрезков в средних классах распределений. Модальный класс распределения по длине трубки составил 19–22 см, а по диаметру – 9–10 мм.
Нарезку стеблей тростника проводили вручную канцелярским ножом для резки бумаги. Так получали отрезки стебля с косым срезом. Отрезки стеблей с прямым срезом получали после резки стеблей тростника пилой-болгаркой. В этом случае для устранения заусенец срезы со стороны входа в трубку обжигали газовой горелкой.
Перед началом сезона гнездования в улей подсаживали пчел в материнских гнездах в виде отдельных пучков трубок, заселенных M. sculpturalis в прошлом сезоне. После того, как пчелы приступали к гнездованию, самок и самцов метили каплями лака для ногтей разного цвета, которые наносили на скутум.

Рис. 5. Гнездовые каналы, прогрызенные в стволе лоха (Elaeagnus angustifolia L.) пчелой-плотником (Xylocopa valga (L.)) и самка Megachile sculpturalis, заселившая брошенное гнездо пчелы-плотника
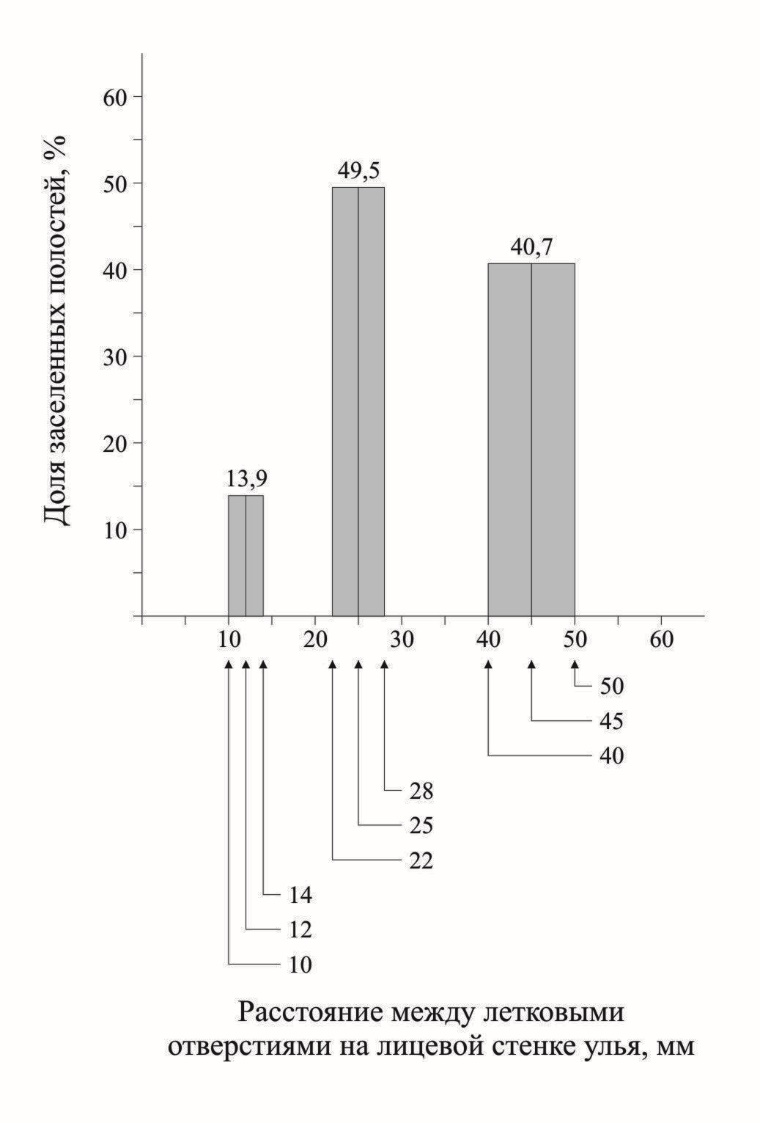
Рис. 6. Процент заселения полостей, расположенных на лицевой стенке улья на разном расстоянии друг от друга
Под шкалой абсцисс указаны средние величины и пределы изменчивости (выделены серым) по каждой группе полостей (n=124).
Экспериментальный улей был установлен на приусадебном участке домостроения одного из районов частной застройки города Симферополя. Наблюдения за гнездованием M. sculpturalis проводили в 2021–2024 годах. Объем исследованного материала приводится в следующем разделе статьи при представлении результатов каждого из проведенных экспериментов. Статистическую обработку данных проводили по общепринятым методикам (Лакин, 1990).
РЕЗУЛЬТАТЫ
Результаты эксперимента по заселению трубок, расположенных на разном расстоянии друг от друга на передней стенке улья, представлены на рисунке 6. Согласно данным рисунка, самки пчел предпочитали заселять трубки, расположенные одиночно на расстоянии 2,5 или 4,5 см друг от друга (49,5 % и 40,7 % заселенных трубок, соответственно). Трубки, собранные в пучки (расстояние между трубками 1,0 см), заселялись менее охотно. При этом, пучки, вставленные в переднюю стенку улья, заселялись охотнее (32,0 % заселенных трубок), чем пучки, расположенные по бокам улья (12,8 % заселенных трубок).
Процент заселения пучков, связанных из трубок с прямым срезом переднего края, оказался равным 33,0 % (n = 124), а с косым – 14,3 % (n = 118), то есть трубки с прямым срезом заселялись чаще в 2,3 раза.
Трубки крайнего внешнего ряда пучка заселялись чаще, чем трубки, расположенные во втором ряду или в центре пучка трубок. Процент заселения трубок внешнего ряда составил 16,5 % (n = 448), второго ряда – 13,5 % (n = 244), а трубок, расположенных в центре пучка – 7, 7 % (n = 52) (рис. 4 в).
Процент заселения трубок, имеющих вогнутое дно составил 46 % (n = 139), а выпуклое – 17 % (n = 93), то есть трубки с ровным и вогнутым дном заселялись в 2,7 раза чаще, чем трубки с неровным и выпуклым дном.
Самки пчел заселяли отрезки тростниковых стеблей диаметром от 7,8 до 15,5 мм (указан внешний диаметр трубок) и длиной от 8 до 32 см. Предпочтение каким-либо трубкам в указанном диапазоне как по диаметру, так и по длине не отмечено (рис. 7).
Выявлена связь между параметрами гнездового канала и возрастом самки. По данным наблюдений за ходом гнездования 4 самок в течение сезона 2022 года каждое следующая трубка, выбираемая ими для строительства гнезда была короче предыдущей и имела меньший диаметр (рис. 8).
ОБСУЖДЕНИЕ
Как отмечено выше, выбирая полость для устройства гнезда, самки M. sculpturalis чаще выбирали трубки, которые располагались на лицевой стенке улья на некотором расстоянии друг от друга, трубки в плотных пучках самки выбирали реже. Следует отметить, что и пучки, и одиночные трубки занимали на передней стенке улья совершенно одинаковое положение, они отличались только расстоянием между отдельными трубками.
И второе – разница между результатами заселения двух вариантов одиночного размещения трубок относительно невелика (49,5 и 40,7 %), в то время как разница между процентом заселения одиночных трубок и трубок в пучках была значительно больше (более 40 % и 13,9 %, соответственно). Кроме того, пучки, вставленные в переднюю стенку улья (расположенные одиночно), так же заселялись чаще, чем пучки, установленные по бокам и под крышей улья и примыкающие друг к другу.
Отмечена и существенная разница в заселении трубок в пределах одного пучка в зависимости от положения трубки относительно его центра. Трубки крайнего внешнего ряда пучка заселялись чаще, чем трубки второго ряда и трубки в центре пучка. При этом важно отметить, что каждая трубка второго ряда и трубка в центре пучка находилась в окружении других трубок, в отличие от трубок первого ряда.
Если выстроить все перечисленные варианты размещения трубок в ряд, в начале которого поместить варианты с самым хорошим заселением, а в конце с самым плохим, то можно заметить, что в этом ряду каждый следующий вариант будет отличаться от предыдущего снижением возможностей для самки запомнить расположение выбранной для гнезда трубки.
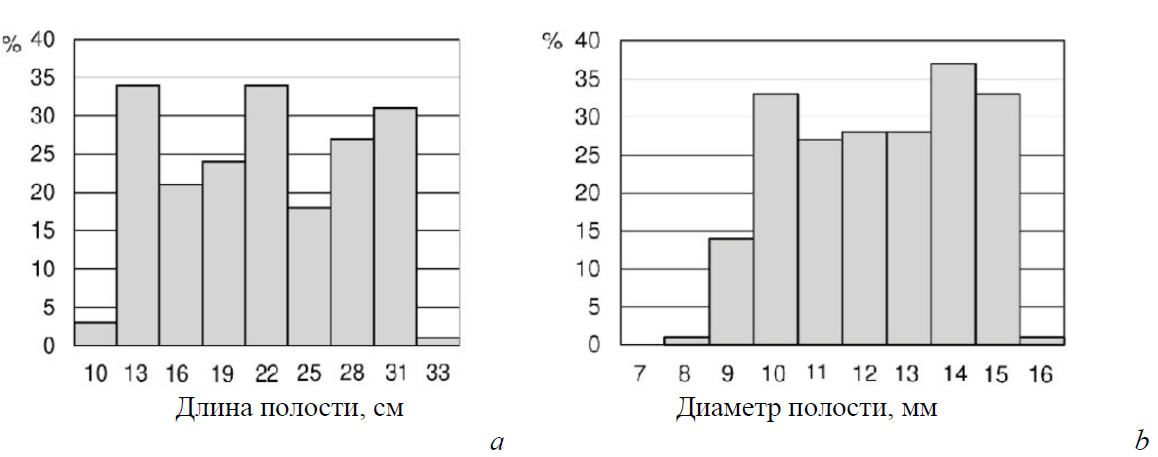
Рис. 7. Гистограммы распределения заселенных самками Megachile sculpturalis полостей по длине (а) и наружному диаметру (b) полости канала (n=313)
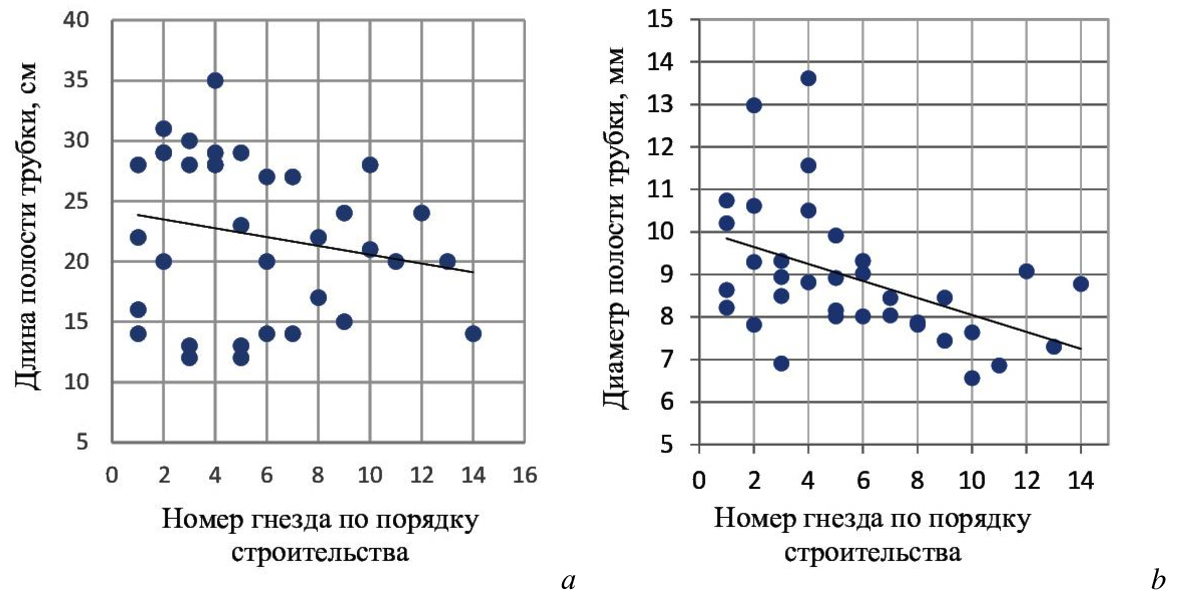
Рис. 8. Связь между возрастом самки Megachile sculpturalis и параметрами выбираемой для строительства гнезда полости
а – связь между возрастом и длиной выбираемой трубки (f=–0,2); b – связь между возрастом и диаметром выбираемой трубки (f=–0,5), (n=35).
В ранее проведенных исследованиях, позволивших провести сравнительный анализ частоты заселения трубок в пучках в зависимости от удаленности их от центра пучка, было показано, что характер заселения рядов трубок у разных видов пчел может существенно отличаться (Жидков, Дубинина, 2014; Иванов и др., 2014). Авторы этих исследований предположили, что частота заселения разных рядов трубок в пучке может определяться не только способностью пчел к ориентации в пределах пучка, но и стремлением самки спрятать гнездо, сделать его менее доступным для паразитов и хищников. При этом, как было показано, возможны 4 варианта сочетания этих двух свойств пчелы и, соответственно, четыре типа распределения заселенных трубок по рядам трубок в пучке. Согласно этой типизации M. sculpturalis можно отнести к видам со средней способностью к запоминанию местоположения выбранной трубки и средне выраженным стремлением сделать гнездо менее заметным.
Более частое заселение пучков, составленных из трубок с прямым срезом переднего края (в сравнении с пучками из трубок с косым срезом), на наш взгляд можно объяснить действием другого фактора. Пучки, составленные из трубок с прямым и косым срезом, выглядят одинаково, если смотреть на них вдоль продольной оси, то есть прямо в торец пучка (рис. 2а). Но если посмотреть на эти два пучка немного сбоку, то картина меняется (рис. 2b). Входные отверстия всех трубок с прямым срезом остаются хорошо заметными, в отличие от трубок с косым срезом, часть входных отверстий которых становится не видна, а часть уже не выглядят как типичные отверстия. Возможно, что именно это делает трубки с косым срезом переднего края менее заметными для пчел при поиске полости для устройства гнезда.
Установленное нами отсутствие у самок M. sculpturalis избирательности в отношении линейных параметров трубки при поиске полости, пригодной для устройства гнезда, является относительно редким явлением среди пчел, поскольку выбор полости оптимальной длины и диаметра у пчел-жильцов имеет важное значение для экономии строительного материала и времени, затраченного на его поиски (Иванов, 1982). Среди исследованных к настоящему времени видов пчел обнаружен только один вид пчел – O. cornuta Latreille, также проявляющий отсутствие избирательности в отношении длины и диаметра выбираемой полости для гнезда (рис. 9 a и b в сравнении с c и d ). Такое исключительное свойство было объяснено (Иванов, 2007, 2010) тем, что данный вид, как отмечал еще Ж. А. Фабр (1898), для устройства своих гнезд при гнездовании в естественных условиях выбирает брошенные гнезда пчел-антофор (Anthophora). В процессе перехода к такому типу гнездования, названному квартиранством (Иванов, 2007 стр. 326), у самок O. cornuta исчезла необходимость оценивать параметры гнездовой полости, в результате чего, инстинкт оценки параметров полости выпал из-под контроля отбора и был постепенно утрачен.
Наблюдения за гнездованием M. sculpturalis показали, что как на территории первичного, так и вторичного ареалов этот вид способен заселять самые разные полости, в том числе и брошенные гнезда других видов пчел (Le Féon, Geslin, 2018). В частности, на территории Северной Америки M. sculpturalis охотно заселяет брошенные гнезда пчел-плотников (Xylocopa). Более того, отмечены случаи агрессивного поведения M. sculpturalis по отношению гнездящимся самкам Xylocopa virginica L. и прямого захвата их гнезд (Laport, Minckley, 2012). По нашим наблюдениям в Крыму самки M. sculpturalis также охотно заселяют брошенные гнезда Xylocopa valga L. (рис. 5b).
Таким образом, можно считать, что выявленное нами отсутствие у самок M. sculpturalis избирательности в отношении длины и диаметра трубки, а также приоритетный выбор самками трубок с вогнутым ровным дном трубок, подобному дну каналов гнезд Xylocopa (рис. 5a), являются следствием проявления склонности M. sculpturalis к заселению чужих гнезд, то есть квартиранству.
Отсутствие избирательности по отношению к диаметру и длине заселяемых каналов установлено нами, видимо, впервые. В известных нам исследованиях Г. Бого с соавторами (Bogo et al., 2024) приводятся данные лишь о диапазоне диаметров, в пределах которого самки M. sculpturalis выбирали гнездовые трубки и их количестве в отдельных классах диапазона. Судить о наличие избирательности без представления о том, сколько трубок было предоставлено пчелам для заселения в каждом классе диапазона диаметров, не представляется возможным.
Выявленное нами уменьшение длины каналов, выбираемых самками для устройства гнезд по мере их старения, можно было бы объяснить постепенной потерей ими волосяного покрова, при участии которого самки оценивают величину диаметра гнездовой трубки, но это предположение противоречит выявленному нами отсутствию у самок стремления оценивать диаметр полости.
Объяснение уменьшения длины полости трубки по причине снижения физической силы, необходимой для оценки длины канала в ходе перемещения самки от входа в полость гнездовой трубки до его дна, не может быть принято по той же причине.
Эти две интересные особенности гнездования M. sculpturalis требует специального экспериментального изучения.
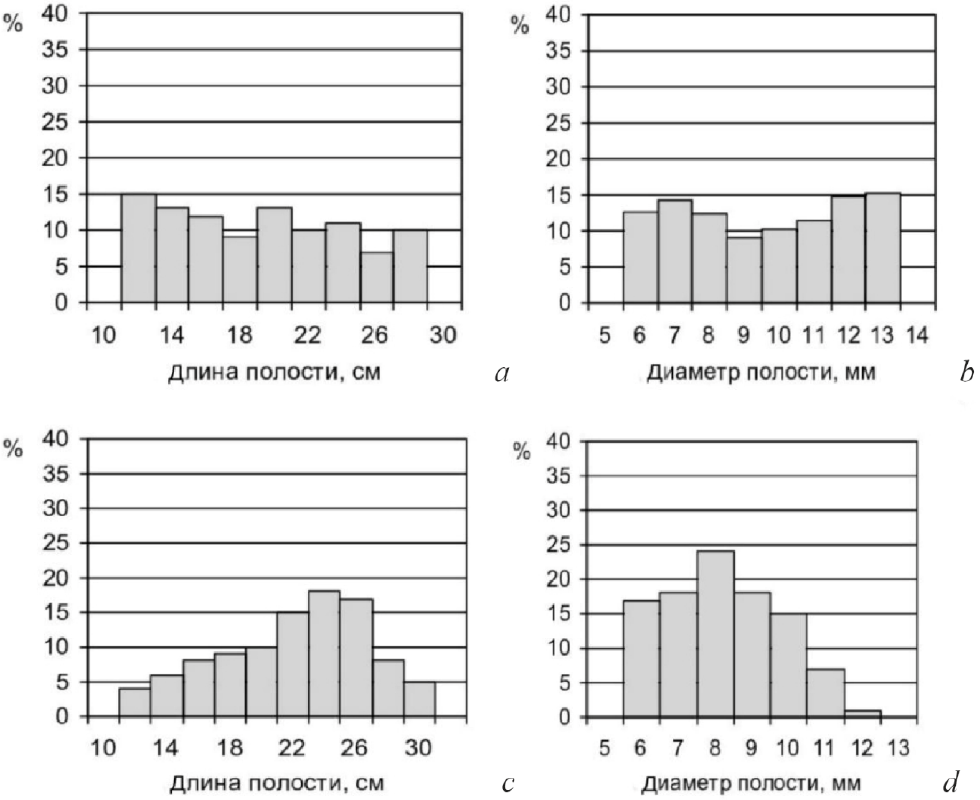
Рис. 9. Гистограммы распределения заселенных полостей по длине (а, c) и диаметру (b, d) для двух видов пчел-мегахилид Osmia cornuta (а, b) Osmia bicornis L. (= rufa) (c, d) (по Иванову, 2007)
ЗАКЛЮЧЕНИЕ
Экспериментальное изучение поведения смоляной пчелы Megachile sculpturalis Smith, 1853 при заселении ульев Фабра, в составе которого пчелам предлагали для заселения отрезки тростниковых стеблей (Phragmites australis) в виде трубок, открытых с одной стороны и закрытых с другой, позволило выявить ряд важных особенностей этологии этого инвазивного вида.
В ходе четырехлетних исследований удалось выявить следующие особенности гнездования M. sculpturalis.
В экспериментальном улье самки пчел предпочитали заселять трубки, расположенные на передней стенке улья одиночно на расстоянии 2,5 или 4,5 см друг от друга (49,5 % и 40,7 % заселенных трубок, соответственно). Трубки, связанные в пучки, заселялись менее охотно. Пучки, вставленные в переднюю стенку улья, заселялись охотнее (32,0 % заселенных трубок), чем пучки, расположенные по бокам и под крышей улья (12,8 % заселенных трубок).
Пучки, собранные из трубок с прямым срезом переднего края, заселялись чаще в 2,3 раза чем пучки, собранные из трубок с косым срезом переднего края.
Трубки крайнего внешнего ряда пучка трубок заселялись чаще, чем трубки, расположенные во втором ряду или в центре пучка трубок. Процент заселения трубок первого ряда составил 16,5 %, второго ряда – 13,5 %, а трубок, расположенных в центре пучка, – 7,7 %.
Относительно низкий процент заселения трубок в отдельных вариантах опыта можно объяснить тем, что в этих вариантах у самок пчел возникали трудности с запоминанием месторасположения выбранной ею для заселения трубки.
Самки пчел заселяли трубки разного диаметра в широком диапазоне от 7 до 16 мм, без предпочтения трубок каких-либо диаметров. Длина заселенных трубок так же широко варьировала от 7 до 35 см. Предпочтения трубок в каком-то диапазоне длины так же не выявлено. Отсутствие предпочтения объясняется потерей самками M. sculpturalis навыков по оценке линейных параметров полости в следствие перехода к квартиранству – заселению гнезд пчел других видов. Это предположение подтверждается известной склонностью M. sculpturalis к заселению гнездовых каналов брошенных гнезд пчел Xylocopa.
Трубки, имеющие вогнутое дно, заселялись в 2,7 раза чаще, чем трубки с выпуклым дном. Эта особенность гнездования M. sculpturalis так же трактуется, как следствие перехода к квартиранству.
Полученные данные важны для понимания успеха в M. sculpturalis как инвазийного вида и могут послужить основой при разработке искусственных конструкций (гостиниц для пчел) для привлечения этого вида к гнездованию в самых разных целях.
- Жидков В. Ю., Дубинина А. В Особенности выбора гнездовых каналов дикими пчелами мегахилидами (Apoidea: Megachilidae) при заселении гнезд-ловушек // Евразийский Союз Ученых. – 2014. – № 9, часть 3. – С. 104–107.
- Иванов С. П. Конструктивные особенности искусственных гнездовий для одиночных пчел // Насекомые-опылители сельскохозяйственных культур (Сб. научн. тр.). – Новосибирск: Сиб. отдел. ВАСХНИЛ, 1982. – С. 79–83.
- Иванов С. П. Биономия пчел-мегахилид (Hymenoptera, Apoidea, Megachilidae) и эволюция их гнездостроительных инстинктов: дисс. … докт. биол. наук: спец. 03.00.24 Энтомология. – Киев: Институт защиты растений, 2007. – 555 с.
- Иванов С. П. Материалы сравнительного изучения строения гнезд пчел-листорезов (Hymenoptera, Megachilidae, Megachile Latr.). Сообщение I. Разнообразие гнездовых построек // Ученые записки Таврического национального университета им. В. И. Вернадского. Серия «Биология, химия». – 2010. – Т. 23, № 3. – С. 68–78.
- Иванов С. П., Жидков В. Ю., Дубинина А. В. Изучение способности пчел-мегахилид (Hymenoptera: Megachilidae) к ориентации по результатам заселения ими гнезд-ловушек // Евразийский Союз Ученых. – 2014. – № 9, часть 3. – С. 44–46.
- Лакин Г. Ф. Биометрия. – М.: Высшая школа, 1990. – 350 с.
- Фабр Ж. А. Инстинкт и нравы насекомых / [Под. ред. И. Я. Шевырева]. – СПб.: Изд-во А. Ф. Маркса, 1898. – 590 с.
- Aguado O., Hernández-Castellano C., Bassols E., Miralles, M., Navarro D., Stefanescu C., Vicens N. (2018). Megachile (Callomegachile) sculpturalis Smith, 1853 (Apoidea: Megachilidae): a new exotic species in the Iberian Peninsula, and some notes about its biology // Butlletí de la Institució Catalana d’Història Natural. – 2018. – Vol. 82. – Р. 157–162.
- Andrieu-Ponel V., Ponel P., Le Féon V., Geslin B., Duvallet G. À propos du comportement de butinage de Megachile sculpturalis Smith, 1853, en France méditerranéenne (Nîmes et Montpellier) (Hymenoptera, Megachilidae) // Bulletin de la Société Entomologique de France. – 2018. – Vol. 123, N 1. – Р. 49–54.
- Bogo G., Fisogni A., Iannone A., Grillenzoni F.-V., Corvucci F., Bortolotti L. Nesting biology and nest structure of the exotic bee Megachile sculpturalis // Bulletin of Entomological Research. – 2024. – Vol. 114, N 1. – Р. 67–76.
- Bila Dubaić J., Raičević J., Plećaš1 M., Lanner J., Nikolić P., Žikić V., Stanisavljević L., Ćetković A. Further range expansion of the sculptured resin bee (Megachile sculpturalis) in serbia and bosnia & Herzegovina // Acta Entomologia Servica. – 2021. – Vol. 26, N 2. – P. 37–63.
- Hinojosa-Díaz I. A. The giant resin bee making its way west: first record in Kansas (Hymenoptera: Megachilidae) // ZooKeys. – 2008. – Vol. 1. – Р. 67–71.
- Ivanov S. P., Fateryga A. V. First record of the invasive giant resin bee Megachile (Callomegachile) sculpturalis Smith, 1853 (Hymenoptera: Megachilidae) in the Crimea // Far Eastern Entomologist. – 2019. – N 395. – P. 7–13.
- Ivanov S. P., Fateryga A. V., Zhidkov V. Yu., Pivovarenko N. A. Giant resin bee Megachile (Callomegachile) sculpturalis Smith, 1853 (Hymenoptera, Apoidea, Megachilidae), an invasive species in the Crimea (notes on its biology) // Ekosistemy. – 2021. – Iss. 28. – P. 122–128.
- Kakutani T., Inoue T., Kato, M., Ichihashi H. Insectflower relationship in the campus of Kyoto University, Kyoto: An overview of the flowering phenology and the seasonal pattern of insect visits // Contributions from the Biological Laboratory, Kyoto. – 1990. – Vol. 27. – Р. 465–521.
- Lanner J., Gstöttenmayer F., Curto M., Geslin B., Huchler K., Orr M. C., Pachinger B., Sedivy C., Meimber H. Evidence for multiple introductions of an invasive wild bee species currently under rapid range expansion in Europe // BMC Ecology and Evolution. – 2021. – Vol. 21. – Article 17.
- Laport R. G., Minckley R. L. Occupation of active Xylocopa virginica nests by the recently invasive Megachile sculpturalis in upstate New York // Journal of the Kansas Entomological Society. – 2012. – Vol. 85, N 4. – Р. 384–386.
- Le Féon V. Geslin B. Écologie et distribution de l’abeille originaire d’Asie Megachile sculpturalis Smith 1853 (Apoidea – Megachilidae – Megachilini): un état des connaissances dix ans après sa première observation en Europe // Osmia. – 2018. – N 7. – Р. 31–39.
- Maeta Y., Kitamura K., Kagino Y., Ikegami N. In-nest behaviors and labor economy of Megachile (Callomegachile) sculpturalis Smith (Hymenoptera, Megachilidae) // Chugoku Kontyo. – 2008. – Vol. 22. – Р. 1–22.
- Mangum W. A., Brooks R. W. First records of Megachile (Callomegachile) sculpturalis Smith (Hymenoptera: Megachilidae) in the Continental United States // Journal of the Kansas Entomological Society. – 1997. – Vol. 70, N 2. – Р. 140–142.
- Parys K., Tripodi A., Sampson B. The giant resin bee, Megachile sculpturalis Smith: new distributional records for the mid- and gulf-south USA // Biodiversity Data Journal. – 2015. – Vol. 3. – Article e6733.
- Quaranta M., Sommaruga A., Balzarini P., Felicioli A. A new species for the bee fauna of Italy: Megachile sculpturalis continues its colonization of Europe // Bulletin of Insectology. – 2014. – Vol. 67. – Р. 287–293.
- Roulston T., Malfi R. Aggressive eviction of the Eastern Carpenter Bee (Xylocopa virginica (Linnaeus)) from its nest by the Giant Resin Bee (Megachile sculpturalis Smith) // Journal of the Kansas Entomological Society. – 2012. – Vol. 85, N 4. – Р. 387–388.
- Westrich P., Knapp A., Berney I. Megachile sculpturalis Smith 1853 (Hymenoptera, Apidae), a new species for the bee fauna of Germany, now north of the Alps // Eucera. – 2015. – N 9. – Р. 3–10.
