
Экосистемы, 32: 42–51 (2022) https://ekosystems.cfuv.ru
![]()
УДК 633.811:674.031.931:581.33:581.143
Никитский ботанический сад – Национальный научный центр РАН Ялта, Республика Крым, Россия
tnkuzmina@rambler.ru
Годичные ритмы жизненных циклов генезиса генеративных структур являются механизмами адаптации растений, позволяющими снизить риски стрессовых воздействий сезонных неблагоприятных абиотических факторов. Jasminum fruticans L. (Oleaceae) – естественный представитель флоры Южного берега Крыма, который цветет в апреле – мае. Генеративные почки у него закладываются летом предшествующего цветению года. Учитывая, что в процессе генезиса пыльники проходит ряд критических периодов, связанных с детерминацией структур, в течение которых они наиболее уязвимы для негативного воздействия стрессовых факторов, то интерес представляет сезонная периодичность генезиса мужской генеративной сферы у данного вида, обусловливающая продуцирование фертильных пыльцевых зерен. В течение 2017–2020 годов определяли длину и стадию развития пыльников длинно- и короткостолбчатых растений. Значение относительной скорости роста пыльника определяли как отношение его прироста за промежуток времени к единице времени. Материал анализировали, начиная от закладки генеративной почки до начала цветения (с июня по апрель). Выделены стадии генезиса пыльников, и показана их линейная очень тесная положительная связь с длиной. Определены периоды усиления и снижения ростовой активности пыльников J. fruticans. Показано, что в течение летнего и осеннего периодов пыльники находятся на стадии покоя. Формирование спорогенной ткани происходит зимой, что сопряжено с усилением интенсивности их роста. Максимальный пик относительной скорости роста пыльников, независимо от морфологической формы растения, происходит в мейотический период. В условиях Южного берега Крыма микроспорогенез у J. fruticans отмечался в первой декаде марта, когда среднесуточная температура находится в пределах +10 °С. Цитоморфологический анализ пыльцевых зерен выявил высокую долю морфологически нормальной пыльцы (70–85 %) у данного вида.
Ключевые слова: пыльник, генезис, микроспорогенез, пыльцевые зерна, сезонная динамика, интенсивность роста, гетеростилия, Jasminum, Oleaceae.
ВВЕДЕНИЕ
Годичные циклы генезиса генеративных структур у покрытосеменных растений обладают адаптивным характером, который обусловливает снижение стрессового воздействия абиотических факторов в критические периоды развития. Для древесных растений областей с умеренным типом климата стратегия генезиса мужской генеративной сферы направлена на снижение рисков негативного влияния низких температур (Миргородская, 2018). В тоже время, для видов, произрастающих в субтропических областях, также важно нивелирование негативного влияния высоких температур и засухи, характерных для продолжительного летнего периода. На Южном берегу Крыма (ЮБК) средиземноморский тип субтропического климата. Зимы на южном побережье короткие и мягкие с преимущественно положительными температурами, а летний период засушливый и жаркий может длиться 4–4,5 месяца (Важов, 1977, Антюфеев, 2014). Для ряда древесных растений Крыма отмечают, что органообразовательные процессы в почках проходят в течение зимнего периода (Методические указания…, 1977). При этом, сведения о сезонной периодичности генезиса генеративных структур древесных растений и кустарников, произрастающих на ЮБК, как правило, основываются на данных фенологических наблюдений и сведений о стадии генезиса органа (Елманов и др., 1969; Галушко, 1980; Яблонский, 1982; Браилко, Кузьмина, 2015). Однако, для определения характера явления целесообразно использование показателей изменения уровня динамического ряда (Куприенко
ISSN 2414-4738 Published by V. I. Vernadsky Crimean Federal University, Simferopol
Сезонная динамика роста и развития пыльников
Jasminum fruticans L. (Oleaceae) в условиях Южного берега Крыма
![]()
- др., 2009). Общепринятым показателем динамики роста органа является его относительная скорость (Лебедев, 1978; Заика, 1982; Шевелуха, 2016). Изучение закономерностей формирование репродуктивных структур растений, как правило, связано с вопросами изучение динамики роста семян и зародышей (Erdelska, 1981; Васильева, Батыгина, 1997). Характеристика роста и развития пыльников, как правило, рассматривается в связи с сопряженностью их размеров и стадий развития (Резникова, 1982; Добровольская и др., 2008). Следует учитывать, что определение интенсивности ростовых процессов позволяет не только выявить закономерности генезиса, но и установить периодичность критических стадий развития, что является одним важных направлений в области репродуктивной биологии растений (Шевченко, Плугатарь, 2019).
Jasminum fruticans L., жасмин кустарниковый, (сем. Oleaceae) представитель флоры Средиземноморья, Средней и Атлантической областей Европы, средней Азии, а также Южного берега Крыма (Голубев, 1996). Особенностью его репродуктивной сферы является диморфная гетеростилия (Guintián et al., 1998; Thompson, Dommée, 2000; Кузьмина, 2017).
Известно, что вид цветет в конце весны, а генеративные почки у него закладываются летом в предшествующий цветению год. Ранее было дано описание генезиса пыльников J. fruticans
(Кузьмина, 2018).
Цель данной работы – дать характеристику сезонной динамики роста пыльников с учетом основных стадий их генезиса у длинно- и короткостолбчатых растений J. fruticans в условиях Южного берега Крыма.
МАТЕРИАЛ И МЕТОДЫ
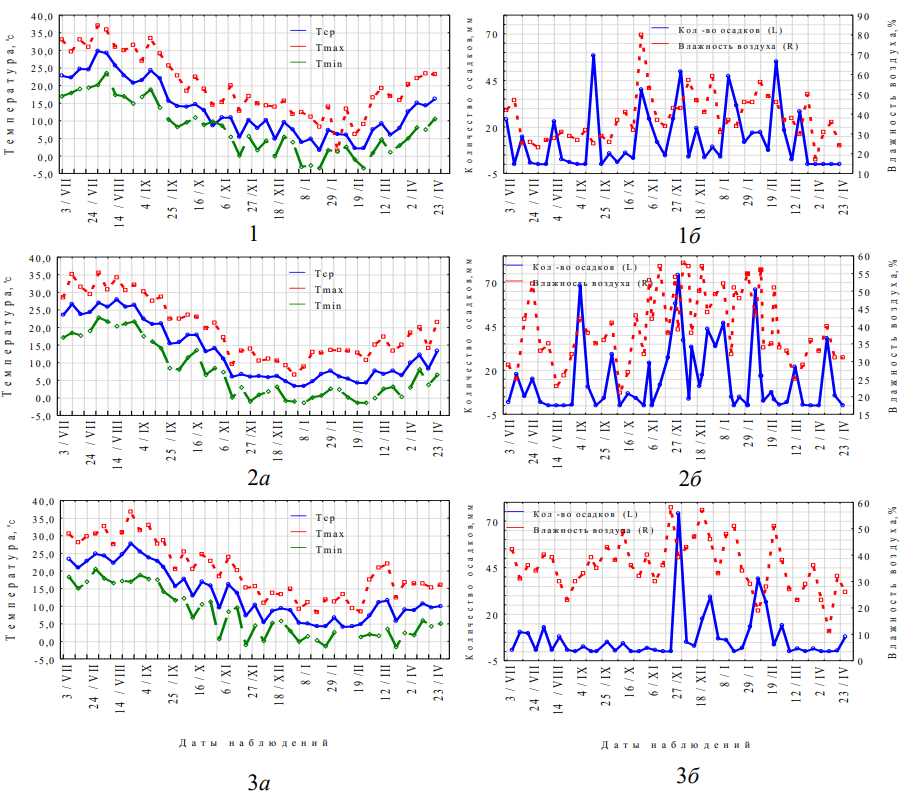
Объектом исследования была мужская генеративная сфера дистильного кустарника флоры Крыма – Jasminum fruticans L., представленного в естественной флоре полуострова особями с цветками двух морфологических форм: длинностолбчатой и короткостолбчатой. Сбор материала проводили на территории Никитского ботанического сада (г. Ялта, Республика Крым). Исследование проходило с июля по апрель в период 2017–2020 годов. На рисунке 1 представлены еженедельные показания средней, максимальной и минимальной температуры воздуха, количества осадков и средней относительной влажности воздуха за 2017–2020 годы в период генезиса пыльников J. fruticans, рассчитанные на основании данных агрометеостанции «Никитский сад» (пгт. Никита, г. Ялта, Республика Крым).
Начиная с момента закладки генеративных почек до начала цветения, еженедельно для каждой морфологической формы анализировали пыльники 18–24 терминальных бутонов. Бутоны фиксировали в течение суток в уксусном алкоголе (alcohol 96 % : acetic acid – 3:1). Временные препараты пыльников окрашивали 1 % ацетоорсеином. Анализ препаратов проводили с помощью микроскопа AxioScope A.1 (Zeiss, Германия). Морфометрические измерения пыльников делали с помощью программного приложения AxioVision Rel. 4.8.2. (Zeiss, Германия).
Для цитоморфологической оценки зрелых пыльцевых зерен для каждой морфологической формы готовили постоянные препараты средних образцов пыльцы из пыльников 50 цветков. Препараты окрашивали метилгрюнпиронином (Шевченко и др., 1986).
- ходе анализа подсчитывали количество морфологически нормальных, аномальных и стерильных пыльцевых зерен. К группе морфологически нормальных были отнесены пыльцевые зерна с однородно окрашенной цитоплазмой и дифференцированными спермиями. При визуальных признаках деструкции цитоплазмы, ядра вегетативной клетки и спермиев пыльцевые зерна оценивались как аномальные. В случае дегенерации содержимого пыльцевого зерна, его характеризовали как стерильное. Подсчет пыльцевых зерен делали с использованием программных приложений AxioVision Rel. 4.8.2 (Zeiss, Германия) и ImageJ 1.48v (National Institutes of Health, США; http:/imagej.nih.gov/ij) в 100 полях зрения.

Данные дескриптивной статистики, корреляционный анализ, построение графиков и расчет относительной скорости роста проводили с помощью программного приложения Statistica 10.0 (StatSoft. Ins., США) (Гашев и др., 2018). Расчет 95 % доверительных интервалов (95 % ДИ) для долей морфологически нормальных, аномальных и стерильных пыльцевых
зерен проводили по методу Уилсона с помощью онлайн-калькулятора (http://vassarstats.net/prop1.html). Морфометрические данные представлены в виде x̅±S x̅, где x̅– среднее арифметическое, S x̅– стандартная ошибка среднего; δ – стандартное отклонение. Статически значимыми приняты значения на уровне p<0,05.
44
Сезонная динамика роста и развития пыльников
Jasminum fruticans L. (Oleaceae) в условиях Южного берега Крыма
![]()
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Согласно общепринятой современной классификации в формировании пыльника выделяют три периода: премейотический, мейотический и постмейотический (Батыгина, Васильева, 2002). Основываясь на характеристиках состояния спорогенной ткани/микроспор или мужского гаметофита, можно выделить 8 стадии развитии пыльника. Для премейотического период характерны стадии археспория и спорогенной ткани. Мейотический период связан с редукционным делением и образованием тетрад микроспор. В постмейотический период проходят стадии молодых микроспор, вакуолизированных микроспор, поздней интерфазы и дифференцирующего деления, двухклеточного пыльцевого зерна и зрелой трехклеточной пыльцы. Размеры пыльников двух морфологических форм J. fruticans с учетом стадии их развития представлены в таблице 1. Корреляционный анализ показал линейную очень тесную положительную зависимость между стадией развития пыльника и его длиной (r=0,98 и r=0,97, соответственно для пыльников длинно- и короткостолбчатых растений). На рисунке 2 представлены фактические кривые длин пыльников длинно- и короткостолбчатых растений J. fruticans.
На основе данных фактических кривых роста были рассчитаны значения относительных скоростей роста (R) (рис. 3), что позволяет проследить периоды повышения и снижения ростовой активности пыльников у длинно- и короткостолбчатых растений J. fruticans. Установлено, что примордии пыльников J. fruticans независимо от морфологической формы растения закладываются в последней декаде июня. Визуально различимыми они становятся в первых числах июля года, предшествующего цветению. Характерно, что в период закладки примордиев пыльников среднесуточная температура воздуха в годы исследования колебалась
- диапазоне +20 – +26 °С, а максимальная температура достигала +35 °С. В целом продолжительность премейотического периода у J. fruticans независимо от морфологической формы растения составляет 8 месяцев. В ходе этого периода развития у J. fruticans можно выделить «первую фазу роста», начало которой приходится на I декаду июля. В I декаде августа она сменяется «фазой покоя», которая длится до II декады декабря. Переход в фазу покоя происходит в наиболее жаркий и засушливый период года, когда, как правило, отсутствуют осадки, а среднесуточная воздуха в этот период достигает +29 °С. Максимальные её значения в отдельные годы наблюдений составляли +36,8 °C. Возобновление ростовых процессов пыльников J. fruticans происходит во I декаде декабря («вторая фаза роста»). В этот период происходит формирование спорогенной ткани и клеточных слоев стенки микроспорангия, которое идет в центробежном направлении, что типично для представителей семейства Oleaceae (Литвиненко, 1987; Камелина, 2009). Характерно, что у J. fruticans, независимо от морфологической формы, эта стадия развития сопровождается усилением интенсивности ростовых процессов. В период стимуляции роста пыльника и образования спорогенной ткани среднесуточные показатели температуры воздуха были в пределах +5 °С.
Максимальный пик ростовой активности соответствует периоду микроспорогенеза и приходится на I–II декады марта, когда среднесуточная температура воздуха находилась в пределах +10 °С. Мейотический период генезиса пыльника J. fruticans самый короткий в ходе формирования пыльника. Показатели относительной скорости роста пыльников на стадии микроспорогенеза составляют 0,07 сут.-1и 0,08 сут.-1, соответственно для пыльников длинно-и короткостолбчатых цветков.
Постмейотический период, в процессе которого происходит созревание микроспорангия и формирования мужского гаметофита, начинается при снижении интенсивности ростовых процессов пыльника, которые сменяются усилением роста. Пики относительной скорости роста приходятся на I–II декады апреля, то есть на период дифференцирующего митоза, двухклеточных пыльцевых зерен и последующего спермиогенного деления, завершающего формирование мужского гаметофита. Именно в постмейотический период становятся четко выраженными различия размеров пыльников у длинно- и короткостолбчатых цветков
45
Кузьмина Т. Н.
![]()
Таблица 1
Длина пыльника на различных стадиях развития
у длинно- (L-форма) и короткостолбчатых (S-форма) растений Jasminum fruticans
| Стадияразвитияпыльника | Длина пыльников, мм | |||||||||||
| L-форма | S-форма | |||||||||||
| Длина | Min | Max | δ | Длина | Min | Max | δ | |||||
| 1 | 0,16 | 0,03 | 0,25 | 0,04 | 0,17 | 0,05 | 0,40 | 0,04 | ||||
| 2 | 0,36 | 0,13 | 0,79 | 0,14 | 0,38 | 0,14 | 0,88 | 0,14 | ||||
| 3 | 0,89 | 0,70 | 1,32 | 0,08 | 0,87 | 0,72 | 1,05 | 0,10 | ||||
| 4 | 1,14 | 0,97 | 1,37 | 0,11 | 1,13 | 0,91 | 1,30 | 0,10 | ||||
| 5 | 1,43 | 1,05 | 1,58 | 0,13 | 1,43 | 1,13 | 1,66 | 0,15 | ||||
| 6 | 1,70 | 1,60 | 1,81 | 0,07 | 1,78 | 1,72 | 1,83 | 0,04 | ||||
| 7 | 1,94 | 1,81 | 2,09 | 0,08 | 1,98 | 1,85 | 2,10 | 0,08 | ||||
| 8 | 2,42 | 2,10 | 2,95 | 0,21 | 2,48 | 2,10 | 3,46 | 0,33 | ||||
| Примечание к таблице. Стадия развития пыльника: 1 – археспориальная ткань; 2 – спорогенная ткань; | |||
| 3 – микроспорогенез; 4 – молодые микроспоры; 5 – вакуолизированные микроспоры; 6 – | |||
| дифференцирующий митоз; 7 – двухклеточные пыльцевые зерна; 8 – трехклеточные пыльцевые зерна. | |||
| Для L-формы n=2043; для S-формы n=2025. | |||
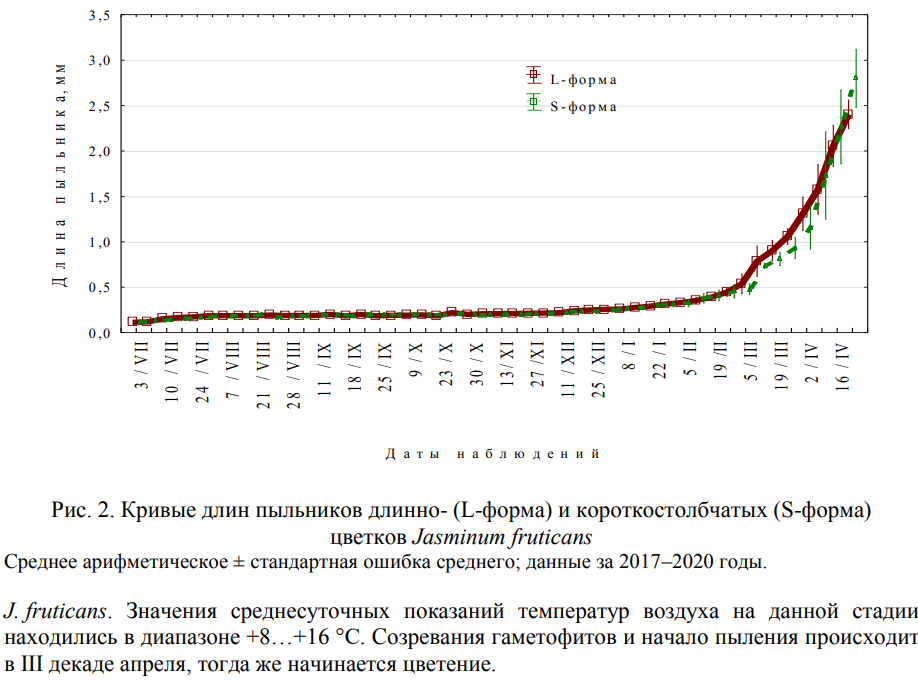
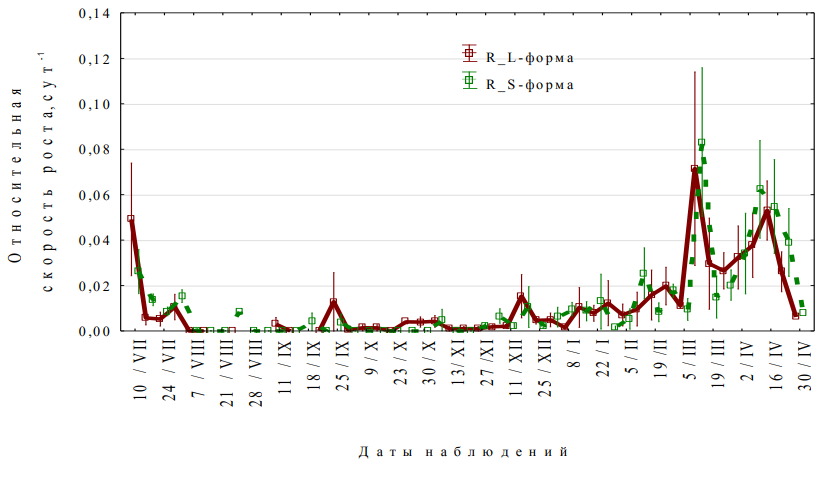
Рис. 3. Динамика средних значений относительной скорости роста пыльников длинно- (L-форма) и короткостолбчатых (S-форма) цветков Jasminum fruticans
Среднее арифметическое ± стандартная ошибка среднего; данные за 2017–2020 годы.
Цитоморфологическая оценка зрелых пыльцевых зерен длинно- и короткостолбчатых цветков J. fruticans показала, что в средних образцах преобладают морфологически нормальные пыльцевые зерна. Так, у длинностолбчатой формы их доля превышает 85 % (95 % ДИ: 85,00‒87,00 %), а в пыльниках короткостолбчатых цветков формируется около 70 % (95 % ДИ: 70,05‒72,64 %) морфологически нормальных пыльцевых зерен (табл. 2), что свидетельствует о высоком уровне фертильности мужского гаметофита.
Таким образом, развитие пыльников J. fruticans, независимо от морфологической формы цветка, характеризуется чередованием периодов усиления и понижения их ростовой активности. Волнообразный характер динамики роста описан для различных структур растений (Яблонский, 1970; Лебедев, 1978; Полевой, Саламатова, 1991; Шевелуха, 2016). При этом изменение интенсивности, локализации и направленности ростовых процессов связывают как с наследственными особенностями, так возрастными и физиолого-биохимическими изменениями, происходящими в растениях. На изменение динамики интенсивности роста оказывают влияние сезонные и суточные колебания факторов внешней среды (температуры, влажности, освещенности, состояние почвенного и воздушного питания) (Лебедев, 1978; Шевелуха, 2016). По мнению Б. П. Токина (1977) внешние факторы могут, как ускорять, так и замедлять и даже приостанавливать развитие.
Известно, что в ходе развития структуры проходят критические периоды. П. Г. Светлов (1960) определил их как периоды детерминации развития, в которые повышается чувствительность клеток к действию внешних факторов, что сопровождается снижением регуляторной деятельности. В ходе органогенеза пыльников критическими считаются ранняя стадия мейоза, стадия тетрад микроспор и стадиях двухклеточных пыльцевых зерен (Добровольская и др., 2009; Batygina, Vasilyeva, 2003). При этом мейотический период, или микроспорогенез, сопровождающийся генетической рекомбинацией и редукцией числа хромосом в ходе образования гаплоидных микроспор, а также качественной перестройкой соматических тканей стенки микроспорангия, является наиболее уязвимым к изменениям влагообеспечения и температурным воздействиям периодом, когда повышаются риски образования аномальных или стерильных гаметофитов (Batygina, Vasilyeva, 2003). Следует
47
Кузьмина Т. Н.
![]()
Таблица 2
Цитоморфологическая характеристика пыльцевых зерен длинно- и короткостолбчатых
цветков Jasminum fruticans
| Длинностолбчатые цветки | Короткостолбчатые цветки | |||||||||
| Доля пыльцевых зерен, % | Доля пыльцевых зерен, % | |||||||||
| Количествопыльцевыхзерен | Количествопыльцевыхзерен | |||||||||
| Год | Морфологическинормальные | Аномальные | Стерильные | Морфологическинормальные | Аномальные | Стерильные | ||||
| 2017 | 884 | 83 | 8 | 9 | 815 | 79 | 7 | 14 | ||
| 2018 | 947 | 85 | 9 | 6 | 771 | 57 | 23 | 20 | ||
| 2019 | 1890 | 87 | 10 | 3 | 1041 | 68 | 24 | 8 | ||
| 2020 | 2157 | 86 | 9 | 5 | 2065 | 75 | 17 | 8 | ||
| Хср±Sx | 5878 | 85,25± | 9± | 5,75± | 4692 | 69,75± | 17,75± | 12,50± | ||
| 0,85 | 0,40 | 1,25 | 4,81 | 3,90 | 2,87 | |||||
отметить, что у J. fruticans микроспорогенез приходятся на I–II декады марта, когда условиях Южного берега Крыма среднесуточная температура воздуха около +10 °C, что существенно снижает риски образование аномалий в ходе редукционного деления клеток, а, следовательно, формирования стерильных гаметофитов.
Учитывая существующую на сегодняшний день типизацию сезонной периодичности основных стадий генезиса мужской генеративной сферы растений, можно отметить, что J. fruticans в большей степени соответствует растениям 1 группы, к которой отнесены представители семейств Caprifoliaceae (Lonicera tolmatchevii Pojark), Grossulariaceae (Ribes nigrum L.), Juglandaceae (Pterocarya pterocarpa (Michx.) Kunth ex l. Iljinsk.), Magnoliaceae (Magnolia sieboldii K. Koch.), Oleaceae (Syringa vulgaris L.) и Rosaceae (Amygdalus ledebouriana Schlecht., Louiseania triloba (Lindl.) Pachom. (Prunus triloba Lindl.), Padus avium Mill., Armeniaca mandschurica (Maxim.) Skvorts; Staphyleaceae (Staphylea colchica Stev.) Миргородская, 2018). Аналогичные ритмы развития пыльников отмечены в условиях Южного берега Крыма для ряда плодовых культур, в частности для персика (Prunus persica (L.) Batshc.), миндаля (Prunus dulcis (Mill.) D.A. Webb.) и абрикоса (Prunus armeniaca L.) (Елманов и др., 1969; Яблонский, 1982). Спорогенную ткань в зимний период отмечают в пыльниках Lonicera tatarica L. (Браилко, Кузьмина, 2015) и культиваров рода Syringa L. (Oleaceae) (Jedrzejuk, Szlachetka, 2005). Известно, что устойчивость спорогенной ткани пыльников обусловлена снижением белоксинтезирующей активности ее клеток (Миргородская, 2018), что служит гарантией стабильности данной структуры к повреждающим воздействиям низких температур (Яблонский, 1970). Однако следует учитывать, что период низких температур, характерный для осенне-зимнего периода, является необходимым условием для полноценных процессов формирования генеративных структур растений (Мирославов, Бармичева, 2009; Котеева и др., 2015; Khodorova et al., 2010; Khodorova, Boitel-Conti, 2013).
48
Сезонная динамика роста и развития пыльников
Jasminum fruticans L. (Oleaceae) в условиях Южного берега Крыма
![]()
ЗАКЛЮЧЕНИЕ
Таким образом, у J. fruticans, независимо от морфологической формы растения, продолжительность генезиса пыльников от закладки до созревания составляет 10 месяцев. Наиболее длительным является премейотический период, который занимает 8 месяцев, в течение которых четыре месяца (с августа по декабрь) пыльник находятся на стадии археспориальной ткани. В этот период практически отсутствует рост пыльников. Усиление интенсивности ростовых процессов начинается с I декады декабря года, предшествующего цветению, на стадии деления и дифференциации клеточных слоев стенки микроспорангия и спорогенной ткани. Переход пыльников к стадии микроспорогенеза отмечается в сезон стабильных положительных температур, который в условиях ЮБК характерен для начала марта. Среднесуточная температура воздуха в этот период находится в пределах +10 °С. Созревание гаметофитов завершается через 1–1,5 месяца после формирования микроспор. Сезонные ритмы генезиса пыльников J. fruticans позволяют нивелировать стрессовые влияния метео-климатических факторов, главным образом, в критические периоды генезиса, что является залогом продуцирования фертильных пыльцевых зерен, доля которых составляет 70–85 %.
Работа выполнена в рамках плановой темы сектора структурной ботаники и репродуктивной биологии растений Никитского ботанического сада – Национального научного центра «Изучение репродуктивных процессов редких и ценных видов семейств Oleaceae, Iridaecae, Asphodelaceae и Campanulaceae с целью познания морфогенетических и функциональных закономерностей системы репродукции и в связи с задачами сохранения и размножения растений», номер государственной регистрации № 0829-2019-0039.
Список литературы
Антюфеев В. В., Казимирова Р. Н., Евтушенко А. П. Агроклиматические, микроклиматические и почвенные условия в приморской полосе Южного берега Крыма. Теоретические основы и практические рекомендации для рационального размещения растений при реконструкции насаждений // Сборник научных трудов ГНБС. – 2014. – Т. 137. – 88 с.
Батыгина Т. Б., Васильева В. Е. Размножение растений: Учебник. – СПб: Изд-во С.-Петербургского университета, 2002. – 232 с.
Браилко В. А., Кузьмина Т. Н. Декоративные жимолости на Южном берегу Крыма: особенности внутрипочечного развития и цветения // Ученые записки Казанского университета. Серия. Естественные науки – Т. 157, кн. 3. – С. 47–57.
Важов В. И. Агроклиматическое районирование Крыма // Труды Никитского ботанического сада. — 1977. – Т. 70. – С. 92–120.
Васильева В. Е., Батыгина Т. Б. Автономность зародыша // Эмбриология цветковых растений. Терминология и концепции. Т. 2. Семя / [Ред. Т. Б. Батыгина]. – СПб: Мир и семья, 1997. – С. 579–588.
Галушко Р. В. Ритмы внутрипочечного развития средиземноморский древесных растений // Сборник научных трудов Никитского ботанического сада. – 1988. – Т. 106. – С. 46–54.
Гашев С. Н., Бетляева Ф. Х., Лупинос М. Ю. Математические методы в биологии. Анализ биологических данных в системе Statistica. – М.: Изд-во Юрайт, 2018. – 208 с.
Голубев В. Н. Биологическая флора Крыма. – Ялта: Изд-во НБС-ННЦ, 1996. – 126 c.
Добровольская А. А., Родионова Г. Б., Воронков А. С., Ковалева Л. В. Спорофитно-гаметофитные взаимодействия в системе пыльник-мужской гаметофит у петунии // Физиология растений. – 2009. – Т. 56, № 3. – С. 437–444.
Елманов С. И., Яблонский Е. А., Шолохов А. М. Анатомо-морфологические и физиологические исследования цветковых почек абрикоса в связи с их зимостойкостью // Труды Государственного Никитского ботанического сада – Т. 60. – 1969. – С. 65–79.
Заика В. Е. Современное состояние теории роста // Математическая биология развития. – М.: Наука, 1982. – С. 40–49.
Камелина О. П. Систематическая эмбриология цветковых растений. Двудольные. – Барнаул: Изд-во «АRTИКА», 2009. – 501 с.
Котеева Н. К., Миргородская О. Е., Булышева М. М., Мирославов Е. А. Формирование пыльцы Ribes nigrum (Grossulariaceae) в связи с периодом пониженных температур // Ботанический журнал. – 2015. – Т. 100, № 10. – С. 1001–1014.
49
Кузьмина Т. Н.
![]()
Кузьмина Т. Н. Диморфизм цветков Jasminum fruticans L. // Вестник Пермского университета. Серия Биология. – 2017. – Вып. 3.– С. 233–239.
Кузьмина Т. Н. Формирование мужских генеративных структур у Jasminum fruticans L. (Oleaceae) // Ботанический журнал. – 2018. – Т. 103, № 5. – С. 654–663.
Куприенко Н. В., Пономарева О. А., Тихонов Д. В. Статистика. Анализ рядов динамики: учебное пособие. –
СПб: Изв-во Политехнического ун-та, 2009. – 203 c.
Лебедев С. И. Физиология растений. – Киев: Вища школа, 1978. – 440 с.
Литвиненко Н. М. Семейство Oleaceae // Сравнительная эмбриология цветковых растений. Davidiaceae – Asteraceae. – Л.: Наука, 1987. – С. 154–158.
Методические указания по фенологическим наблюдениям над деревьями и кустарниками при их интродукции на юге СССР / [Сост. Голубева И. В., Галушко P. В., Кормилицын А. М.]. – Ялта, 1977. – 25с.
Миргородская О. Е. Развитие мужского гаметофита некоторых древесных покрытосеменных растений в условиях умеренного климата: автореф. дис. … канд. биол. наук. – СПб: Ботанический института им. В. Л. Комарова РАН, 2018. – 22 с.
Мирославов Е. А., Бармичева Е. М. Апоптозоподобная деградация пыльцевых зерен у Scilla sibirica связана с отсутствием пониженных температур при их развитии // Физиология растений – 2009. – Т. 56, № 6. – С. 942–947.
Полевой В. В., Саламатова Т. С. Физиология роста и развития растений: учебное пособие. – Л: Изд-во Ленингр. ун-та, 1991. – 240 с.
Резникова С. А. Цитология и физиология развивающегося пыльника. – М.: Наука, 1984. – 272 с.
Светлов П. Г. Теория критических периодов развития и ее значение для понимания принципов действия среды на онтогенез // Вопросы цитологии и общей физиологии / [Ред. Ю. И. Полянский]. – М.–Л., 1960. – С. 263– 285.
Токин Б. П. Общая эмбриология. Учебник для биологических специальностей университетов. – М.: Высшая школа, 1977. – 512 с.
Шевелуха В. С. Рост растений и его регуляция в онтогенезе. Избранные сочинения. Т. 1. – М.: Изд-во ИТРК, 2016. – 594 с.
Шевченко С. В., Ругузов И. А., Ефремова Л. М. Методика окраски постоянных препаратов метиловым зеленым и пиронином // Бюллетень Государственного Никитского ботанического сада. – 1986. – Т. 60. – С. 99–101.
Шевченко С. В., Плугатарь Ю. В. Репродуктивная биология цветковых растений и задачи рационального приропользования // Plant Biology and Horticulture: theory, innovation. – 2019. – № 3 (152). – С. 15–25.
Яблонский Е. А. Влияние температуры на зимнее развитие генеративных почек абрикоса // Физиология растений. – 1982. – Т. 29, №. 6. – С. 1075–1082.
Яблонский Е. А. Темпы роста и зимостойкость сортов абрикоса, персика и миндаля // Исследования по физиологии, биохимии, цитологии, эмбриологии и радиобиологии растений. Труды Государственного Никитского ботанического сада. – 1970. – Т. 44. – С. 50–61.
Batygina T. B., Vasilyeva V. E. Periodization in the development of flowering plant reproductive structures: critical periods // Acta Biologica Cracoviensia. Series Botanica. – 2003. – Vol. 45, N 1. – P. 27–36.
Erdelska O. Dynamics of early embryo development in reference to endosperm and embryo types in angiosperm // Acta Soc. Bot. Polon. – 1981. – Vol. 50, N 1–2. – P. 269 – 272.
Guitián J., Guitián P., Medrano M. Floral biology of the distylous Mediteranean shrub Jasminum fruticans (Oleaceae) // Nordic Journal of Botany. – 1998. – Vol. 18. – P. 195–201.
Jedrzejuk A., Szlachetka W. Development of flower organs in common liliac (Syringa vulgaris L.) cv. Mme Florent Stepman //Acta Biologica Cracoviensla. Series Botanica. – 2005. – Vol. 42, N 2. – P. 41–52.
Khodorova N. V., Boitel-Conti M. The role of temperature in the growth and flowering of geophytes // Plants. – 2013. – Vol. 2. – P. 699–711.
Khodorova N. V., Miroslavov E. A, Shavarda A. L., Laberche J.-C., Boitel-Conti M. Bud development in corydalis (Corydalis bracteata) requires low temperature: a study of developmental and carbohydrate changes // Annals of Botany. – 2010. – Vol. 105. – P. 891–903.
Thompson J. D., Dommée B. Morph-specific patterns of variation in stigma height in natural populations of distylous Jasminum fruticans. // New Phytol. – 2000. – Vol. 148. – P. 303–314.
Vasilyeva V. E., Batygina T. B., Titova G. E. Morpho-physiological correlation in the development of the reproductive structures of Nelumbo nucifera Gaertn. // Phytomorphology: An International Journal of Plant Morphology. – 1987. – Vol. 37. – P. 349–357.
50
Сезонная динамика роста и развития пыльников
Jasminum fruticans L. (Oleaceae) в условиях Южного берега Крыма
![]()
Kuzmina T. N. Seasonal dynamics of growth and development of anthers of Jasminum fruticans L. (Oleaceae) in conditions of the Southern Coast of Crimea // Ekosistemy. 2022. Iss. 32. P. 42–51.
The annual rhythms of life cycles of the genesis of generative structures are the mechanism of plant adaptation. Jasminum fruticans L. (Oleaceae) is a representative of natural flora of the Southern Coast of Crimea. The species is blooming in April–May. Generative buds are formed in summer in the year preceding flowering. The study is aimed at revealing the seasonal periodicity of the genesis of the male generative sphere of J. fruticans. The length and the stage of development of anthers of long- and short-styled plants were determined during 2017–2020.The value of the relative growth rate (R) of the anther was calculated as the ratio of its growth over a certain period of time to a unit of time.The material was takenfor the analysis, starting with the formation of the generative bud and up to the beginning of flowering (from June to April).The stages of the genesis of anthers were determined. A linear close positive relationship between the stage of anther development and its length was shown. The stages of increase and decrease of growth activity of anthers were revealed.The anthers of J. fruticans were dormant during summer and autumn periods. Growth increased in winter, when sporogenic tissue was formed. The maximum peak of the relative growth rate of anthers occured during the meiotic period, regardless of the morphological form of the plant. In the conditions of the Southern Coast of Crimea, microsporogenesis of J. fruticans was observed in the first decade of March, when the average daily temperature was within +10 °C. Cytomorphological analysis of pollen grains revealed a high proportion of morphologically normal pollen (70–85 %).
Key words: anther, genesis, microsporogenesis, pollen grains, seasonal dynamics, growth intensity, heterostyly, Jasminum, Oleaceae.
Поступила в редакцию 18.02.22
Принята к печати 12.12.22
51
