
Экосистемы, 33: 119–133 (2023) https://ekosystems.cfuv.ru
![]()
УДК 582.594.2:[581.46+581.5] (477.75)
Сволынский А. Д., Иванов С. П., Курамова В. В.
Крымский федеральный университет имени В. И. Вернадского Симферополь, Республика Крым, Россия svolinskiy@gmail.com; spi2006@list.ru, viktoriya.bekirova@ya.ru
Проанализировано распространение орхидеи пальчатокоренника римского (Dactylorhiza romana) на Крымском полуострове. На южном берегу Крыма (ЮБК) вид встречается на протяжении всего западного южнобережья и только в двух пунктах на востоке восточного южнобережья (Карадаг). В зоне горных лесов Крыма D. romana встречается только в западных частях южного и северного макросклонов крымских гор, при этом на северном макросклоне этот вид поднимается до высоты 1100 м н.у.м. В предгорной зоне Крыма D. romana встречается только на границе с зоной горных лесов – в трех отдельных локалитетах в западной части предгорий,
- одном – в центральной части и в трех – на крайнем востоке этой природной зоны. Фенодаты периода цветения колеблются в зависимости от погодных условий сезона. Начало цветения – с 5 по 10 апреля, окончание – 10–20 мая. Фенодаты цветения южнобережных и горно-лесных ценопопуляций совпадают, при этом сумма положительных температур начала цветения ценопопуляций на ЮБК составляет 650 °С, а в горных лесах – 200 °С. Распределение генеративных особей в двух местообитаниях в горных лесах Крыма (Осиновая балка) и на ЮБК (гора Кастель) крайне неравномерно – от плотных скоплений, в которых расстояние между цветущими особями измеряется 1–50 см, до разреженных с расстоянием между особями 1–3 м и более. Такое распределение сильно отклоняется от нормального и представляет собой распределение «разломанного стержня» МакАртура. Южнобережная и горно-лесная ценопопуляции D. romana достоверно отличаются по длине цветоносов, соцветий и количеству цветков в соцветиях. Генеративные особи на горе Кастель по всем перечисленным параметрам превосходят экземпляры из Осиновой балки. Растения из южнобережного локалитета так же характеризуются бо́льшими величинами коэффициента вариации по большинству параметров. Существенные отличия в сумме положительных температур, определяющих фенодаты периода цветения, а также морфологические отличия растений южнобережной и горно-лесной ценопопуляций трактуются как наличие в Крыму двух экологических форм D. romana.
Ключевые слова: Dactylorhiza romana, Orchidaceae, распространение в Крыму, пространственное распределение особей в ценопопуляции, фенология цветения, морфометрические показатели соцветий, полуостров Крым.
ВВЕДЕНИЕ
Орхидеи (Orchidaceae Juss.) широко распространены по всем континентам Земли (кроме Антарктиды) и обладают исключительным разнообразием форм. По оценкам разных авторов это семейство насчитывает от 25 до 30 тысяч видов (Dressler, 1994; Delforge, 2006; Christenhusz, Byng, 2016).
Орхидные – одно из молодых, эволюционно продвинутых семейств цветковых растений. Видам этого семейства присуща высокая степень стенотопности и сложность биоценотических связей, особенно ярко выраженная в разнообразии взаимоотношений с опылителями (Pijl, Dodson, 1966; Фегри, Пейл, 1982; Claessens, Kleynen, 2011; Ackerman et al., 2023; и др.). Изучение сложных биоценотических связей орхидей важно для разработки мер охраны этих в своем большинстве редких растений. Из 45 видов орхидей, занесенных в Красную книгу Республики Крым, 10 видов оценены как сокращающиеся в численности, а 5 видов – как исчезающие (Красная книга…, 2015).
Изучение экологии орхидей Крыма проводится по целому ряду направлений, важнейшие из которых – репродуктивная биология (Курамова и др., 2020; Лагутова и др., 1996; Вахрушева, Кучер, 1997; Назаров, 2016, 1996, 1995 и др.), возрастная и пространственная
ISSN 2414-4738 Published by V. I. Vernadsky Crimean Federal University, Simferopol
Сволынский А. Д., Иванов С. П., Курамова В. В.
![]()
структура ценопопуляций (Попкова, 2009; Кобечинская и др., 2012; Сволынский и др., 2014а; и др.) и взаимоотношения с опылителями, в частности, выявление их видового состава и эффективности опыления (Назаров, Иванов, 1990; Назаров, Ефетов, 1993; Холодов и др., 2002; Иванов и др., 2010а, 2010б; Фатерыга, Иванов, 2012; Сволынский и др., 2014б; Lagutova, Chebotar, 1990; Nazarov, 1995; и др. ).
Природа взаимодействия с опылителями и особенности жизнедеятельности орхидеи пальчатокоренника римского (Dactylorhiza romana (Sebast.) Soó) в пределах его обширного ареала изучены фрагментарно (Claessens, Kleynen, 2011; Хомутовский, 2015; Ефимов, 2022).
- пределах Крыма ранее фрагментарно изучены репродуктивная биология вида, распространение по полуострову и фенология цветения (Лагутова и др., 1996; Кучер и др., 2013, 2016; Иванов, Сволынский, 2015; Фатерыга и др., 2019).
Цель наших исследований – провести анализ распространения пальчатокоренника римского в Крыму, изучить фенологию цветения, пространственное распределение цветущих особей в отдельных ценопопуляциях, выявить морфометрические показатели генеративных особей, произрастающих на южном берегу Крыма и в горных лесах в сравнительном аспекте.
МАТЕРИАЛЫ И МЕТОДЫ
Антэкологию D. romana исследовали в двух ценопопуляциях: первая располагалась на горе Кастель (100 м н. у. м.) на склоне северо-западной экспозиции в типичном для южного берега Крыма шибляке из дуба пушистого (рис. 1a); вторая – на западном склоне горного массива Северная Демерджи в Осиновой балке (700 м н. у. м.), под пологом леса из дуба скального (рис. 2a).
Основной материал для исследований был собран в сезоны 2013–2015 годов, дополнительный – в последующие годы.
Анализ распространения D. romana в Крыму проводили в основном по литературным источникам и результатам собственных наблюдений авторов.
Пространственное распределение цветущих особей в ценопопуляциях интересовало нас, прежде всего, с позиции взаимоотношений D. romana с опылителями, перемещающимися от одного соцветия к другому. Поэтому выявление общего характера распределения генеративных особей в пространстве (тип дисперсии) проводили по Ю. Одуму (1975, стр. 265–267) в интерпретации Л. Дайса (Dice, 1952). Количественную оценку степени сгруппированности особей осуществляли с использованием метрического классификатора, известного как метод «ближайшего соседа» (Харитонов, 2005). Метод основан на последовательном измерении расстояний между цветущими особями. При этом в качестве следующей особи каждый раз выбирали ближайшую, исключая уже учтенные. Результаты промеров представляли в виде гистограммы распределения расстояний между цветущими особями. Более детальное описание данной методики приведено в работе В. В. Курамовой с соавторами (2022).
Фенологию цветения изучали по данным периодических (с интервалом в 6–10 дней) посещений изучаемых ценопопуляций, в ходе которых осуществляли просмотр соцветий, сопровождавшийся подсчетом бутонов, распустившихся и отцветших цветков не менее чем на 25 соцветиях, что позволило выявить динамику распускания и увядания цветков, сроки начала и окончания цветения, а также среднюю продолжительность цветения одного цветка.
Влияние погодных условий на сроки цветения изучали по результатам расчета суммы активных температур, накопленных к той или иной фенодате, суммируя положительные температуры воздуха превышающие 5 °С.
- исследовании учитывались климатические данные с интернет-ресурса http://www.pogodaiklimat.ru по четырем метеостанциям Крыма (Ангарский перевал (Синоптический индекс (СИ) – 33958), Алушта (СИ – 33959), Ялта (СИ – 33990), Симферополь (СИ – 33946)).
120
Особенности антэкологии Dactylorhiza romana (Orchidaceae) в Крыму:
распространение, фенология, пространственное размещение и морфометрия цветущих растений

Рис. 1. Орхидея Dactylorhiza romana (южный берег Крыма, г. Кастель)
Участок леса на склоне горы Кастель – место произрастания D. romana на южном берегу Крыма (а).
Соцветия орхидеи с белыми (b), желтыми (с) и пурпурными (d) цветками.
Морфометрию особей проводили в полевых условиях, без изъятия растений из природы. Длину и диаметр окружности соцветий измеряли у особей с полностью распустившимися цветками. Диаметр окружности соцветий (диаметр горизонтальной проекции соцветия) измерялся для последующего расчета периметра окружности соцветия и плотности цветков в соцветии. Общая высота растений измерялась от поверхности почвы до верхушки соцветия. Выборка из южнобережной ценопопуляции включала растения с разной окраской цветков (рис. 1 b, с, d).
121
Сволынский А. Д., Иванов С. П., Курамова В. В.

Рис. 2. Орхидея Dactylorhiza romana (горный Крым, урочище Осиновая балка)
Участок леса в урочище Осиновая балка – место произрастания D. romana в горных лесах Крыма (а).
Плотная группа цветущих растений (б) и особь, цветущая одиночно (с).
Все количественные данные обрабатывались в программе Microsoft Office Excel 2010. Величину и характер корреляционной связи оценивали по Н. И. Черновой (1999). Асимметрию распределений оценивали по формуле Пирсона:
![]() ,
,
где: ![]() – средняя; Мо – мода; σ – сигма.
– средняя; Мо – мода; σ – сигма.
122
Особенности антэкологии Dactylorhiza romana (Orchidaceae) в Крыму:
распространение, фенология, пространственное размещение и морфометрия цветущих растений
![]()
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Распространение пальчатокоренника римского в Крыму. Распространение орхидеи D. romana на Крымском полуострове носит своеобразный характер. На южном берегу Крыма вид встречается в основном в западной части этой природной зоны. В восточной части этой природной зоны вид обнаружен только в двух пунктах – на самом востоке зоны в районе Карадага. На горных лугах яйл не обнаружен.
-
- зоне горных лесов Крыма D. romana встречается только в западных частях южного и северного макросклонов, при этом на северном макросклоне этот вид поднимаемся – до 1100
- н.у.м. и местами спускается до границ с зоной предгорий. В зоне предгорий Крыма D. romana встречается только на границе с зоной горных лесов – в трех локалитетах в западной части предгорий, в одном – в центральной части и в трех – на крайнем востоке.
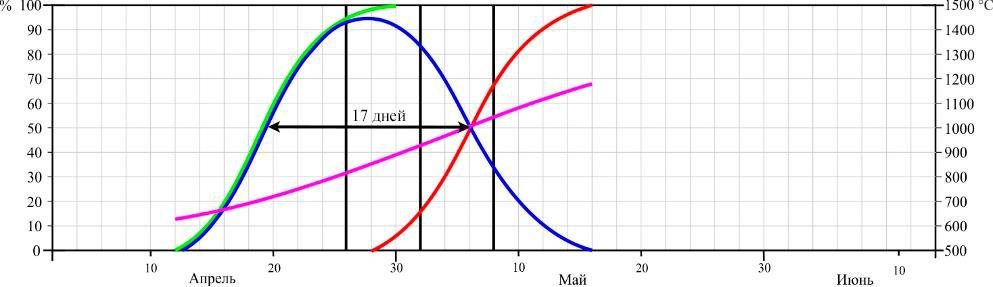
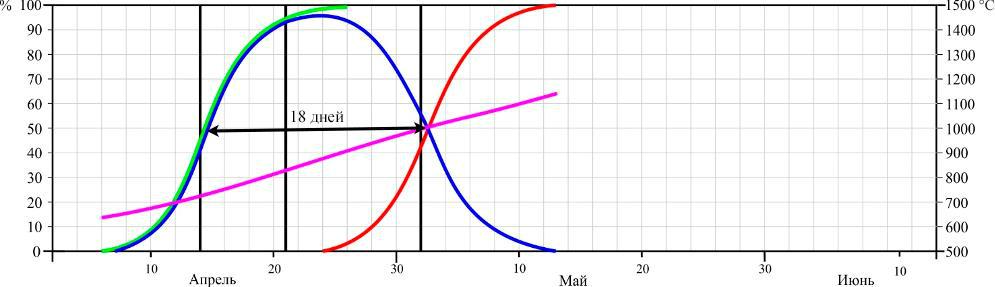
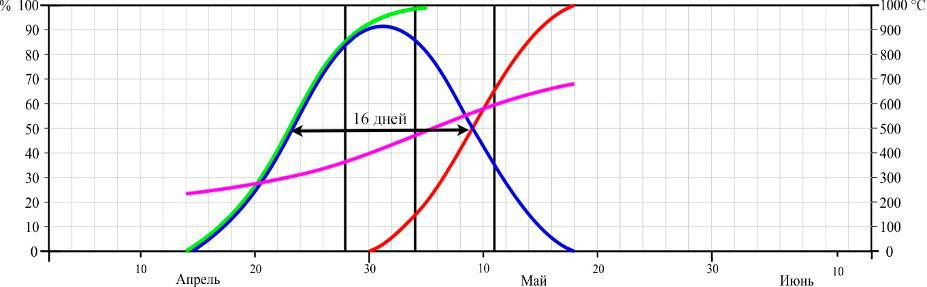
Фенология цветения. Динамика цветения D. romana в двух пунктах произрастания в сезоны 2013–2015 годов представлена на рисунках 3 и 4.
2013 г.
2014 г.
2015 г.
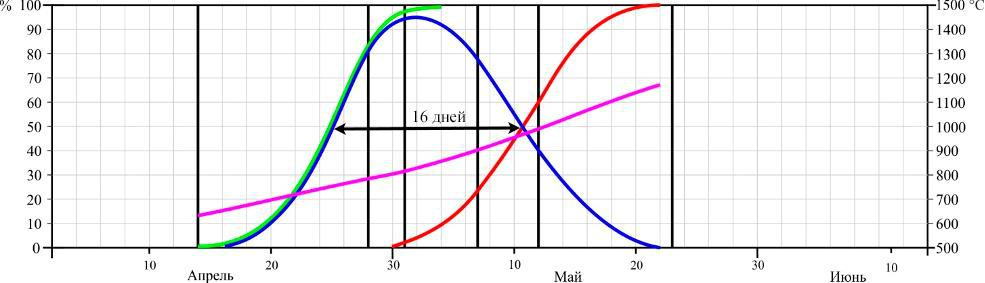
Рис. 3. Динамика цветения Dactylorhiza romana на горе Кастель в разные годы (по Иванов, Сволынский, 2015 с изменениями)
- – доля цветущих цветков от их общего количества; ▬ – кумулята накопления распустившихся цветков; ▬ – кумулята накопления отцветших цветков, ▬ – кумулята накопления положительных температур. Даты учета соотношения бутонов, цветущих и отцветших цветков отмечены вертикальными линиями.
123
Сволынский А. Д., Иванов С. П., Курамова В. В.
![]()
2013 г.
2014 г.
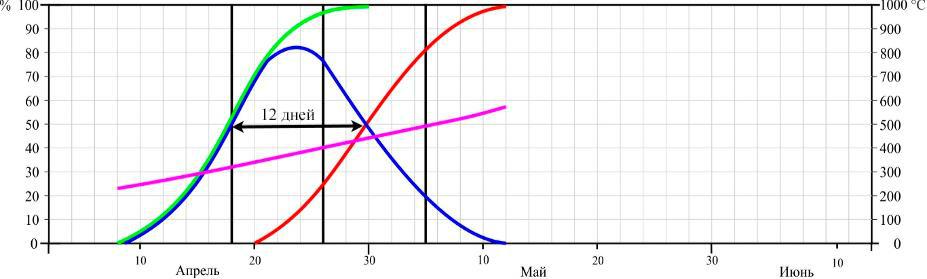
2015 г.
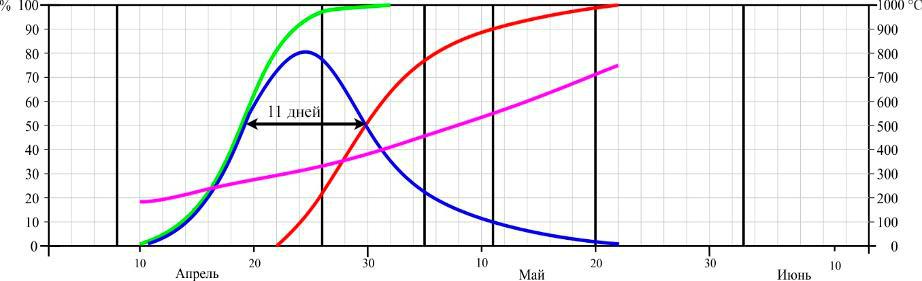
Рис. 4. Динамика цветения Dactylorhiza romana в Осиновой балке в разные годы (по Иванов, Сволынский, 2015 с изменениями)
- – доля цветущих цветков от их общего количества; ▬ – кумулята накопления распустившихся цветков; ▬ – кумулята накопления отцветших цветков, ▬ – кумулята накопления положительных температур. Даты учета соотношения бутонов, цветущих и отцветших цветков отмечены вертикальными линиями.
Из данных рисунков следует, что начало цветения D. romana существенно не отличается между двумя пунктами, если сравнивать их по отдельным годам. Так в 2013 году на горе Кастель начало цветения выпало на 12 апреля, в 2014 – на 8 апреля, и в 2015 году – на 16 апреля, а в Осиновой балке – на 18, 12, 22 апреля соответственно. Еще в большей мере это относится и к датам конца цветения: 16, 12, 22 мая на горе Кастель и 18, 12, 22 мая в Осиновой балке по тем же годам соответственно.
Таким образом, по наблюдениям за 3 сезона, фенодаты начала цветения ценопопуляций D. romana на южном берегу Крыма и в горном районе колебались в пределах 9 и 4 дней соответственно. Даты окончания цветения – в пределах 8 и 10 дней соответственно. При этом даты как начала, так и окончания цветения практически полностью совпадали по отдельным годам.
124
Особенности антэкологии Dactylorhiza romana (Orchidaceae) в Крыму:
распространение, фенология, пространственное размещение и морфометрия цветущих растений
![]()
Продолжительность цветения D. romana на горе Кастель в 2013 году составила 34 дня, а пик цветения наблюдался с 24–30 по апреля; в 2014 году продолжительность цветения – 32 дня с пиком цветения в период с 20 по 27 апреля; в 2015 году продолжительность – 37 дней, пик цветения – с 29 апреля по 5 мая. В Осиновой балке продолжительность цветения в 2013 году – 34 дня, пик цветения – с 28 апреля по 4 мая; в 2014 году продолжительность цветения – 32 дня, пик цветения – с 21 по 26 апреля; в 2015 году продолжительность цветения – 36 дней, пик цветения – с 22 по 27 апреля.
Таким образом, и по фенодатам начала и конца цветения, и по продолжительности периода цветения наблюдалось крайне близкое сходство южнобережной ценопопуляции и ценопопуляции, произрастающей в горно-лесной зоне.
Продолжительность цветения одного цветка на горе Кастель в середине периода цветения составила 17 дней в 2013 году, 18 – в 2014 и 16 – в 2015 году. В среднем за 3 года – 17 дней.Продолжительность цветения одного цветка в горно-лесной зоне Крыма составила 16 дней в 2013 году, 12 – в 2014 и 11 – в 2015 году. В среднем за 3 года – 13 дней.
-
- ходе трехлетних исследований было установлено, что сумма положительных температур начала цветения на горе Кастель в 2013 году составила 623 °С, в 2014 – 641 °С и 634 °С в 2015 году, а окончания цветения – 1179 °С, 1187 °С, 1152 °С соответственно годам.
- Осиновой балке сумма положительных температур начала цветения по 2013, 2014 и 2015 годам составила 241 °С, 233 °С и 187 °С, а окончания – 679 °С, 571 °С и 714 °С соответственно.
Пространственное распределение генеративных особей в пределах отдельных ценопопуляций. О характере размещения генеративных особей D. romana в отдельных ценопопуляциях дает представление рисунок 5.
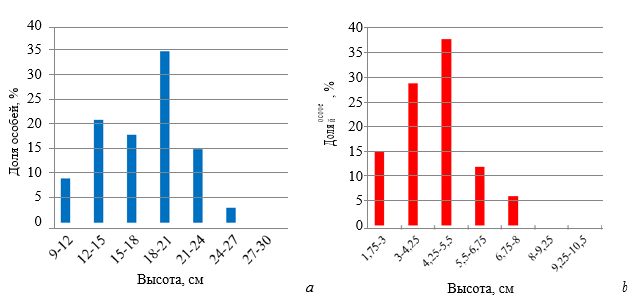
Рис. 5. Гистограммы распределения цветущих растений Dactylorhiza romana по расстоянию до ближайшего соседа на горе Кастель (а) и в Осиновой балке (b)
Сходство гистограмм распределения цветущих растений по расстоянию до ближайшего соседа в южнобережной и горно-лесной ценопопуляциях, свидетельствует, что пространственная структура генеративных особей D. romana в данных локалитетах аналогична. Как на горе Кастель, так и в Осиновой балке преобладающее число особей (более 70 %) располагалось на расстоянии от 1 до 50 см друг от друга. В следующих классовых интервалах распределения наблюдали последовательное убывание числа особей. Такой тип крайне неравномерного дискретного распределения носит название – распределение «разломанного стержня» МакАртура (MacArthur, 1957) и отражает ситуацию, когда особи со средним значением показателя не составляют большинство и не представляют модальный класс, как при нормальном распределении, известном как распределение Гаусса.
125
Сволынский А. Д., Иванов С. П., Курамова В. В.
![]()
Необходимо отметить, что распределение особей на горе Кастель все же несколько отличалось от распределения особей в Осиновой балке. В интервале от 0 до 50 см на горе Кастель большинство особей располагалось на расстоянии менее 25 см друг от друга, а в Осиновой балке – более 25 см. Эта особенность не отражена на рисунке, поскольку эти различия проявляются в пределах одного класса распределения.
Морфометрические показатели генеративных особей. Значения основных морфометрических показателей цветущих растений D. romana в двух изученных ценопопуляциях представлены в таблице 1.
Таблица 1
Значения основных морфометрических показателей цветущих растений Dactylorhiza romana
в двух пунктах произрастания в Крыму (по Сволынскому, 2016 с изменениями)
| Пункт | Параметр | Показатели выборки и величины параметров | |||||
| n | min–max | ±σ | σ2 | CV, % | |||
| Общая высота растения, | 50 | 9,1–32,4 | 19,3±4,7 | 22,1 | 24 | ||
| см | |||||||
| Длина цветоноса, см | 50 | 6,4–20,3 | 13,9±3,2 | 10,4 | 23 | ||
| Гора | Длина соцветия, см | 50 | 2,4–12,1 | 5,3±1,8 | 3,5 | 35 | |
| Периметр окружности | 50 | 5,9–14,1 | 10,6±1,6 | 2,6 | 15 | ||
| Кастель | |||||||
| соцветия, см | |||||||
| Число цветков в | 50 | 4–30 | 12,7±5,3 | 28,7 | 42 | ||
| соцветии | |||||||
| Плотность цветков в | 4,4* | ||||||
| соцветии шт./см2 | |||||||
| Общая высота растения, | 48 | 9,1–24,3 | 16,2±3,3 | 10,9 | 20 | ||
| см | |||||||
| Длина цветоноса, см | 48 | 5,9–18,7 | 12,1±2,8 | 8,1 | 24 | ||
| Осиновая | Длина соцветия, см | 48 | 2,3–6,0 | 4,2±0,9 | 0,8 | 21 | |
| Периметр окружности | 48 | 5,6–12,5 | 9,3±1,7 | 2,8 | 18 | ||
| балка | |||||||
| соцветия, см | |||||||
| Число цветков в | 48 | 5–19 | 9,9±2,6 | 6,9 | 27 | ||
| соцветии | |||||||
| Плотность цветков в | 4,2* | ||||||
| соцветии шт./см2 | |||||||
![]()
Примечание к таблице. Статистически значимые различия (p<0,05) между параметрами растений из двух разных пунктов зарегистрированы по всем показателям, кроме отмеченных звездочкой (*).
Как следует из данных таблицы, генеративные особи на горе Кастель по всем параметрам превосходят экземпляры из Осиновой балки, в том числе и по трем коэффициентам вариабельности из пяти. Наименьшие значения вариации, в обоих пунктах произрастания, характерны для параметра «периметр окружности соцветия», а наибольшие – для параметра «количество цветков в соцветии».
На рисунке 6а представлены гистограммы распределения растений по общей высоте из двух изученных ценопопуляций. Гистограмма распределения особей по общей высоте растений для ценопопуляции на горе Кастель соответствует нормальному распределению, но
- небольшой отрицательной асимметрией (Аs=−0,15) – сдвигом значений в большую сторону. Здесь наиболее часто встречались растения высотой 21–24 см. Гистограмма распределения особей по общей высоте цветущих растений в Осиновой балке так же соответствует нормальному распределению. В данной ценопопуляции растения модального класса меньше по высоте – 18–21 см.
126
Особенности антэкологии Dactylorhiza romana (Orchidaceae) в Крыму:
распространение, фенология, пространственное размещение и морфометрия цветущих растений
![]()
Рис. 6. Гистограммы распределения цветущих растений по общей высоте (а) и длине соцветия (b) в двух локалитетах Dactylorhiza romana
Без штриховки – ценопопуляция в Осиновой балке; штриховка – ценопопуляция на горе Кастель.
Сравнивая гистограммы распределения особей по длине соцветия (рис. 6b), следует отметить, что модальный интервал распределения в Осиновой балке – 3–4,25 см, а на горе Кастель – 4,25–5,5 см. На горе Кастель в крайний справа по шкале класс включены растения
- длиной соцветия в пределах 9,25–10,5 см, при этом отмечен один исключительный экземпляр с длиной соцветия 12,4 см, который не был включен в рисунок. Максимальная длина соцветий D. romana в Осиновой балке достигала всего 6,5 см. Кроме того, гистограмма распределения растений горной ценопопуляции по длине соцветия имеет четко выраженную правостороннюю асимметрию (Аs=0,66) – сдвиг в сторону меньших значений.
- обоих локалитетах выявлена сильная положительная связь ( r=0,96 и 0,97) между показателями: общая высота растения и длина цветоноса (табл. 2), но слабая связь – между длиной цветоноса и длиной соцветия, особенно низкая в горной ценопопуляции (r=0,39). Это позволяет предположить, что длина цветоноса и длина соцветия у D. romana определяются разными факторами.
Таблица 2
Связь между некоторыми параметрами генеративных особей Dactylorhiza romana
- ценопопуляциях на южном берегу Крыма (г. Кастель)
- в горно-лесной зоне (Осиновая балка)
| Пункт | Параметр | Общая | Длина | Длина | Число | |
| высота растения | цветоноса | соцветия | цветков | |||
| Общая высота растения | 1 | |||||
| Гора Кастель | Длина цветоноса | 0,96 | 1 | |||
| Длина соцветия | 0,87 | 0,69 | 1 | |||
| Число цветков | 0,73 | 0,58 | 0,85 | 1 | ||
| Общая высота растения | 1 | |||||
| Осиновая | Длина цветоноса | 0,97 | 1 | |||
| балка | Длина соцветия | 0,61 | 0,39 | 1 | ||
| Число цветков | 0,96 | 0,94 | 0,58 | 1 | ||
| 127 |
Сволынский А. Д., Иванов С. П., Курамова В. В.
![]()
На рисунке 7 представлены ранжированные ряды цветущих растений D. romana по общей высоте, длине цветоноса и соцветия. По этим рядам наглядно видны отличия растений из разных ценопопуляций по размерным показателям, большее или меньшее соответствие нормальному распределению, а также вариабельность параметров.
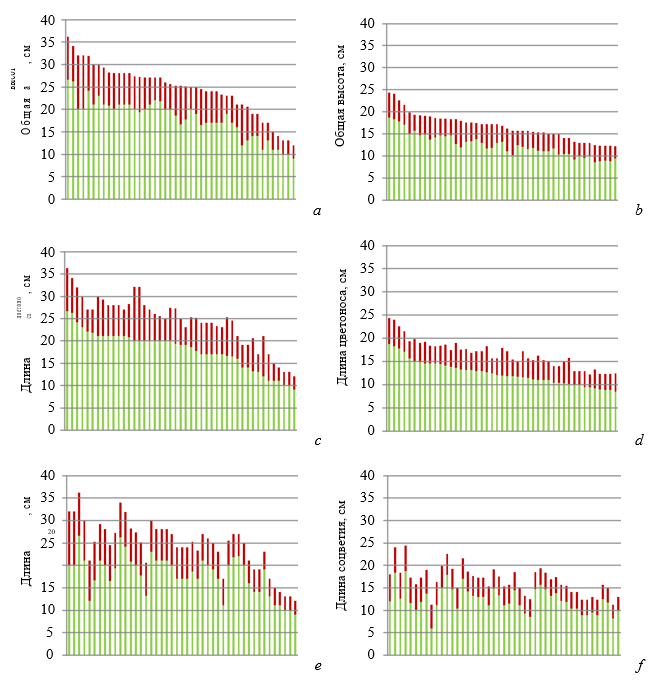
Рис. 7. Ряды растений Dactylorhiza romana ранжированные по общей высоте цветущего растения (a и b), по длине цветоноса (c и d) и длине соцветия (e и f)
Зеленая часть столбцов – цветонос, красная – соцветие; a, c, e – гора Кастель; b, d, f – Осиновая балка.
- частности, ряды особей, ранжированные по общей высоте растения (рис. 7 а, b) и длине цветоноса (рис. 7 с, d), демонстрируют большее соответствие нормальному распределению особей из южнобережной ценопопуляции и большую их вариабельность.
128
Особенности антэкологии Dactylorhiza romana (Orchidaceae) в Крыму:
распространение, фенология, пространственное размещение и морфометрия цветущих растений
![]()
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Первые сведения о распространении орхидей в пределах Крымского полуострова с указанием расположения отдельных ценопопуляций были представлены Е. В. Вульфом (1930). В дальнейшем сведения о распространении отдельных видов орхидей в Крыму были приведены в Красной книге Украины (Червона книга…, 2009) и Красной книге Республики Крым (Красная книга…, 2015), а затем были существенно дополнены в двух книгах (Kreutz et al., 2018; Фатерыга и др., 2019). Сравнение характера распространения D. romana в Крыму с другими видами орхидей показывает, что распространение этой орхидеи в Крыму полностью не повторяет ни один из крымских видов этого семейства. Определенное сходство с распространением D. romana по Крыму демонстрирует только Anacamptis coriophora (L.) R. M. Bateman, Pridgeon & M. W. Chase и в какой-то мере Neottia ovata (L.) Bluff & Fingerh и Orchis mascula (L.) L. Для выявления причин сходства и отличий в характере распространения отдельных видов орхидей по Крыму необходимо проведение специальных исследований.
Размещение генеративных особей в пределах отдельных ценопопуляций у разных видов может существенно отличаться. С некоторыми ограничениями характер размещения особей
- пределах местообитания, видимо, можно рассматривать как видовой признак. В то же время степень и формы проявления дискретности или континуальности отдельных ценопопуляций могут изменяться в течении последовательного ряда лет в ответ на воздействие как благоприятных, так и неблагоприятных факторов среды (Заугольнова, 1976; Фардеева и др., 2009; Фардеева, 2018). Анализ ряда методик оценки дискретности особей в отдельных ценопопуляциях (Фардеева, 2018) показал, что каждая из них может быть успешной в зависимости от особенности объектов изучения и целей исследований. Использованный нами метод «ближайшего соседа», не требующий применения сложного математического аппарата и формализованный в виде распределения МакАртура, обеспечил однозначность интерпретации результатов наших исследований. В частности, в соответствии с результатами исследований М. Б. Фардеевой (2018), характер выявленного нами пространственного распределения особей можно считать свидетельством относительного благополучного состояния как южнобережной, так и горно-лесной ценопопуляций D. romana. Случайное размещение особей, согласно данным М. Б. Фардеевой, указывает на угнетенное состояние ценопопуляции, а дискретное – признак благополучия.
Ранее такой тип распределения был отмечен у Orchis provincialis Balb. ex Lam. & DC. (Сволынский и др., 2014а), однако авторы не распознали его как особый тип распределения, и приняли за некое отклонение от нормального.
Представленные в данной публикации данные по фенологии цветения D. romana касаются трех отдельных сезонов и не противоречат ранее известным сведениям по данному виду (Фатерыга и др., 2019). Новизна и ценность полученных нами данных заключается в том, что они включают сведения о сумме температур наступления отдельных фенодат периода цветения D. romana, что дает возможность прогнозировать сроки наступление отдельных фаз цветения данного вида в любом из текущих сезонов.
Кроме того, периодический подсчет соотношения цветков разного состояния (бутоны, цветущие, отцветшие) позволил выявить динамику зацветания и динамику отцветания соцветий, а также среднюю продолжительность цветения одного цветка в каждом из локалитетов и в каждом сезоне отдельно.
-
- неожиданным результатам наших исследований можно отнести тот факт, что продолжительность цветения одного цветка в ценопопуляции в горно-лесной зоне оказалась меньше, чем на южном берегу Крыма. В среднем за три года наблюдений в горной ценопопуляции этот показатель составил 14 дней, а в южнобережной – 17.
Следует отметить, что продолжительность цветения отдельного цветка у орхидей разных видов может сильно варьировать – от одного дня до нескольких десятков дней (Sugiura et al., 2001; Huda, Wilcock, 2012; Вахрамеева и др., 2014). При этом, было установлено, что на продолжительность цветения цветка влияет наличие в цветке поллинариев. Среднее время цветения одного цветка после удаления поллинариев меньше, чем у цветка с неудалёнными
129
Сволынский А. Д., Иванов С. П., Курамова В. В.
![]()
поллинариями (Zhang et al., 2014). Таким образом, продолжительность цветения одного цветка, зависит от интенсивности опыления орхидей, то есть численности пчел-опылителей.
Исходя из этого, можно предположить, что интенсивность опыления оказывает влияние
- на общую положительность периода цветения орхидеи. Возможно, что именно этим можно объяснить выявленный нами больший разброс значения суммы температур даты окончания периода цветения (Δ = 40 °С) по сравнению с разбросом значения суммы температур даты начала цветения (Δ = 25 °С). Более детальный анализ влияния интенсивности опыления на динамику цветения изученных ценопопуляций D. romana будет представлен в следующей публикации, полностью посвященной вопросам опыления.
Особый интерес вызывают полученные нами сведения о совпадении сроков начала цветения в горной и южнобережной ценопопуляциях D. romana при значительной разнице в сумме положительных температур этой фенодаты. Возможно, что совпадение сроков цветения южнобережных и горных ценопопуляций D. romana сформировалось постепенно в ходе проникновения этого вида из южнобережья в горные районы Крыма под действием отбора, поддерживающего особи наиболее ранних сроков цветения. Такие особи лучше опылялись, поскольку использовали привычный комплекс опылителей, и соответственно были более успешны в размноженнии. В результате этих процессов в Крыму могли образоваться две экологические формы D. romana.
Приведенные нами данные о наличии статистически значимых отличий между величинами ряда морфометрических показателей цветущих растений D. romana из южнобережной и горной ценопопуляций согласуются с известными ранее аналогичными данными, включая отличия по вегетативным органам (Сволынский, 2016), а также данными А. В. Фатерыги и С. А. Й. Кройца (Fateryga, Kreutz, 2014), которые обратили внимание на то, что произрастающие на южном берегу растения D. romana отличаются от растений из других районов Крыма несколько более широкими листьями и несколько более крупными цветками. Кроме того, известно, что D. romana в Крыму представлена двумя морфологическими формами, отличающимися окраской цветков. Еще Е. В. Вульф (1930) писал, что «В Крыму встречается одновременно разновидности с желтыми и фиолетовыми цветами». Позже было установлено, что растения с фиолетовыми цветками встречаются только на южном берегу Крыма (Fateryga, Kreutz, 2014; Красная книга…, 2015; Kreutz et al., 2018).
Таким образом, полученные нами данные о разной сумме положительных температур основных фенодат периода цветения южнобережной и горной ценопопуляций D. romana, при совпадении основных фенодат этого периода, наличие достоверных морфологических и некоторых других отличий генеративных растений этих ценопопуляций, позволяют предположить существование в Крыму как минимум двух экологических форм D. romana.
Дальнейшие исследования, включая изучение ценопопуляций D. romana, расположенных в нижнем поясе северного макросклона крымских гор и в предгорьях, позволят внести большую ясность в эти вопросы.
ВЫВОДЫ
- Распространение пальчатокоренника римского в Крыму достаточно своеобразно и не повторяет картину распространения ни одного из крымских видов орхидей. Определенное сходство в распространения с D. romana по Крыму демонстрирует только
Anacamptis coriophora (L.) R. M. Bateman, Pridgeon & M. W. Chase и в какой-то мере Neottia ovata (L.) Bluff & Fingerh и Orchis mascula (L.) L.
- По наблюдениям в течении 3-х последовательных сезонов, фенодаты начала и окончания цветения ценопопуляций D. romana на южном берегу Крыма и в горном районе колебались в пределах 4–10 дней, при этом практически полностью совпадали по отдельным годам. Продолжительность цветения исследованных ценопопуляций варьировала от 34 до 42 дней и так же существенно не отличалась при сравнении по отдельным годам.
- Продолжительность цветения одного цветка варьировала в горно-лесной ценопопуляции по годам от 11 до 16 дней (в среднем 14), а в южнобережной – от 16 до 18 (в
130
Особенности антэкологии Dactylorhiza romana (Orchidaceae) в Крыму:
распространение, фенология, пространственное размещение и морфометрия цветущих растений
![]()
среднем 17).
- В ходе трехлетних исследований было установлено, что сумма положительных температур начала цветения на горе Кастель по наблюдениям 2013–2015 годов колебалась от 623 до 641 °С, а в Осиновой балке – от 187 до 241 °С. Причины столь существенной разницы сумм положительных температур между южнобережной и горно-лесной ценопопуляциями при равенстве основных фенодат цветения требуют специального изучения.
- Пространственное распределение цветущих растений D. romana в пределах ценопопуляции соответствует распределению «разломанного стержня» МакАртура. Как в южнобережных, так и горно-лесных ценопопуляциях плотные группы особей чередуются с одиночно цветущими растениями. Более 70 % особей располагаются на расстоянии от 1 до 50 см друг от друга. Выявленная дискретность распределения генеративных особей трактуется как свидетельство относительного благополучия изученных ценопопуляций.
- Южнобережная и горно-лесная ценопопуляции D. romana достоверно отличаются по длине цветоносов, соцветий и числу цветков в соцветиях. Генеративные особи на горе Кастель (южный берег Крыма) по всем перечисленным параметрам превосходят экземпляры из Осиновой балки (горно-лесная зона Крыма).
- Растения из южнобережной ценопопуляции, характеризуются бо́льшими величинами коэффициента вариации, чем растения из горно-лесной. Наименьшие значения вариации в обоих пунктах произрастания отмечены для параметра «периметр окружности соцветия», а наибольшие – для «количество цветков в соцветии».
- Анализ морфометрических показателей цветущих растений показал сильную положительную связь между высотой растения и длиной цветоноса и слабую связь между длиной цветоноса и длиной соцветия, особенно низкую в горной ценопопуляции (r=0,39), что позволяет предположить, что длина соцветий D. romana не определяется факторами, которые влияют на размеры вегетативных частей растений.
- Некоторые особенности распространения пальчатокоренника римского на полуострове, существенные отличия в сумме положительных температур, определяющих фенодаты периода цветения при совпадении значений основных фенодат, а также ряд морфологических отличий растений южнобережной и горной ценопопуляций дают основания предположить наличие в Крыму двух экологических форм D. romana.
Список литературы
Вахрамеева М. Г., Варлыгина Т. И., Татаренко И. В. Орхидные России (биология, экология и охрана). – М.:
Товарищество научных изданий КМК, 2014. – 437 с.
Вахрамеева М. Г., Денисова Л. В., Никитина С. В., Самсонов С. К. Орхидеи нашей страны. – М.: Наука, 1991. – 224 с.
Вахрушева Л. П., Кучер Е. Н. Особенности репродуктивного усилия некоторых видов орхидей Крыма // Экосистемы Крыма, их оптимизация и охрана. – 1997. – Вып. 9. – С. 65–67.
Вульф Е. В. Флора Крыма. Том 1, выпуск 3. – Л.: Изд. Никитского Ботанического Сада, 1930. – 126 с. Заугольнова Л. Б. Неоднородность строения ценопопуляций во времени и пространстве // Ботанический
журнал, 1976. – Т. 61, № 2. – С. 187–196.
Иванов С. П., Фатерыга А. В., Тягнирядно В. В. Сравнительная оценка эффективности опыления орхидей в урочище Аян // Бюллетень Государственного Никитского ботанического сада. – 2008. – Вып. 97. – С. 10–14.
Иванов С. П., Фатерыга А. В., Тягнирядно В. В. Эффективность опыления орхидей (Orchidaceae), цветущих одиночно и группами // Бюллетень Государственного Никитского ботанического сада. – 2009а. – Вып. 98. – С. 22– 26.
Иванов С. П., Холодов В. В., Фатерыга А. В. Орхидеи Крыма: состав опылителей, разнообразие систем и способов опыления и их эффективность // Ученые записки Таврического национального университета имени В. И. Вернадского. Сер. «Биология, химия». – 2009б. – Т. 22, № 1. – С. 24–34.
Иванов, С. П., Сволынский А. Д. Особенности фенологии цветения ранневесенних меллитофильных орхидей (Orchidacea) в Крыму // Экосистемы. – 2015. – Вып. 1. – С. 85–96.
Кобечинская В. Г., Отурина И. П., Сверкунова Н. В. Динамика развития и цветения орхидеи кокушника комарникового (Gymnadenia conopsea) в Крыму // Экосистемы, их оптимизация и охрана. – 2012. – Вып. 6. – С. 151–161.
131
Сволынский А. Д., Иванов С. П., Курамова В. В.
![]()
Красная книга Республики Крым. Растения, водоросли и грибы / [Ред. А. В. Ена, А. В. Фатерыга]. – Симферополь: ИТ «АРИАЛ», 2015. – 480 с.
Курамова В. В., Иванов С. П., Сволынский А. Д. Некоторые антэкологические особенности орхидеи Neotinea tridentata в Крыму: пространственное размещение, параметры и цветовая гамма соцветий // Экосистемы. – 2022. – Вып. 31. – P. 143–154.
Лагутова О. Н., Назаров В. В., Шевченко С. В. Семенное воспроизведение Dactylorhiza romana (Orchidaceae) в Крыму // Ботанический журнал. – 1996. – Т. 81, № 5. – С. 59–69.
Назаров В. В., Ефетов К. А. Участие пестрянок (Lepidoptera, Zygaenidae) Крыма в опылении орхидеи Anacamptis pyramidalis (Orchidaceae) // Зоологический журнал. – 1993. – Т. 72, № 10. – С. 54–67.
Назаров В. В., Иванов С. П. Участие пчел рода Chelostoma Latr. (Hymenoptera, Megachilidae) в опылении мимикрирующих видов Cephalanthera rubra (Z.) Rich. и Campanula taurica Juz. в Крыму // Энтомологическое обозрение – 1990. – Т. 69, № 3. – С. 534–537.
Назаров В. В. Репродуктивная биология орхидных Крыма: дис. … канд. биол. наук: 03.00.05 Ботаника. – СПб.:
Ботанический институт им. В. Л. Комарова, 1995. – 123 с.
Одум Ю. Основы экологии. – М.: Мир, 1975. – 740 с.
Попкова Л. Л. Состояние и сохранение популяций влаголюбивых видов орхидных Крыма // Заповедники Крыма. Теория, практика и перспективы заповедного дела в Черноморском регионе: V Международная научно-практическая конференция, 22–23 октября 2009 г. – Симферополь, 2009. – С. 208–213.
Сволынский А. Д., Иванов С. П., Фатерыга А. В. Особенности антэкологии ятрышника прованского (Orchis provincialis, Orchidaceae) в Крыму: фенология, пространственное распределение, морфометрия цветков и соцветий
- Экосистемы, их оптимизация и охрана. – 2014а. – Вып. 10. – С. 68–76.
Сволынский А. Д., Иванов С. П., Фатерыга А.В. Особенности антэкологии ятрышника прованского (Orchis provincialis, Orchidaceae) в Крыму: опылители, система их привлечения, уровень опыления // Экосистемы, их оптимизация и охрана. – 2014б. – Вып. 11. – С. 144–157.
Сволынский А. Д. Антэкология четырех видов ранневесенних энтомофильных орхидей (Orchidaceae Juss.) Крыма: дис. … канд. биол. наук: 03.02.08 Экология. – Ялта: Никитский ботанический сад, 2016. – 160 с.
Фардеева М. Б. Пространственная неоднородность популяций клубнеобразующих орхидей и методы ее изучения на примере Neottianthe сucullata (L.) Schlechter // Экосистемы. – 2018. – Вып. 16 (46). – С. 75–85.
Фардеева М. Б., Чижикова Н. А., Бирючевская Н. В., Рогова Т. В., Савельев А. А. Математические подходы
- анализу пространственно-возрастной структуры популяций дерновинных видов трав // Экология. – 2009. – №
4. – С. 249–257.
Фатерыга А. В., Ефимов П. Г., Свирин С. А. Орхидеи Крымского полуострова. – Симферополь: ИТ
«АРИАЛ», 2019. – 224 с.
Фатерыга А. В., Иванов С. П. Экология опыления видов рода Epipactis (Orchidaceae) в Крыму // Экосистемы, их оптимизация и охрана. – 2012. – Вып. 6. – С. 136–150.
Фегри К., Пэйл Л. Основы экологии опыления. – М.: Мир, 1982. – 381 с.
Харитонов С. П. Метод «ближайшего соседа» для математической оценки распределения биологических объектов на плоскости и на линии // Вестник Нижегородского университета. Серия биология. – 2005. – № 1. – С.
213–221.
Холодов В. В., Назаров В. В., Иванов С. П. Видовой состав пчел (Hymenoptera, Apoidea) опылителей некоторых видов орхидей в Крыму // Экосистемы Крыма их оптимизация и охрана. – 1998. – Вып. 10. – С. 81–86.
Холодов В. В., Назаров В. В., Иванов С. П. Насекомые посетители и опылители орхидеи Orchis purpurea Huds. (Orchidaceae) в Крыму // Экосистемы Крыма, их оптимизация и охрана. – 2002. – Вып. 12. – С. 77–80.
Червона книга України. Рослинний світ / [Ред. Я. П. Дідух]. – К.: Глобалконсалтинг, 2009. – 912 с.
Чернова Н. И. Математическая статистика. – Новосибирск: Новосибирский государственный университет,
2007. – 148 с.
Ackerman J. D., Phillips R. D., Tremblay R. L., Karremans A., Reiter N., Peter C., Bogarín D., Pérez-Escobar O. A., Liu H. Beyond the various contrivances by which orchids are pollinated: global patterns in orchid pollination biology // Botanical Journal of the Linnean Society. – 2023. – P. 1–30.
Christenhusz M. J. M., Byng J. W. The number of known plants species in the world and its annual increase // Phytotaxa. – 2016. – Vol. 261, N 3. – P. 201–217.
Claessens J., Kleynen J. The Flower of the European Orchid. Form and Function. – Voerendaal, 2011. – 439 p.
Delforge P. Orchidées d’Europe, d’Afrique du Nord et du Proche-Orient. Quatrième édition. – Paris: Delachaux et Nistlé, 2016. – 544 p.
Dice L. R. Measure of spaсing between individuals within a population // Contributions from the Laboratory of Vertebrate Biology, University of Michigan. – 1952. – Vol. 55. – P. 1–23.
Dressler R. Phylogeny and Classification of the Orchid Family. – Cambridge University Press, 1994. – 314 p.
Fateryga A. V., Kreutz C. A. J. Checklist of the orchids of the Crimea (Orchidaceae) // Journal Europäischer
Orchideen. – 2014. – Bd. 46, Heft 2. – S. 407–436.
Huda M. K., Wilcock С. С. Rapid floral senescence following male function and breeding systems of some tropical orchids // Plant Biology. – 2012. – Vol. 14, N 2. – P. 278–284.
Kreutz C. A. J., Fateryga A. V., Ivanov S. P. Orchids of the Crimea. – Sint Geertruid: Kreutz Publishers, 2018. – 576 p.
132
Особенности антэкологии Dactylorhiza romana (Orchidaceae) в Крыму:
распространение, фенология, пространственное размещение и морфометрия цветущих растений
![]()
Lagutova O. I., Chebotar A. A. Pollination ecology of Orchis provincalis Bald. // Embryology and Seed Reproduction: XI International Symposium, July 3–7 1990: Abstracts of the reports. – St. Petersburg, 1990. – P. 306–307.
MacArthur R. H. On the relative abundance of bird species // Proceedings of the National Academy of Sciences of the United States of America. – 1957. – Vol. 45, N 3. – P. 293–295.
Nazarov V. V. Pollination of Steveniella satyrioides (Orchidaceae) by wasps (Hymenoptera, Vespoidea) in the Crimea // Lindleyana. – 1995. – Vol. 95, N 2. – P. 109–144.
Sugiura N., Fujie T., Inoue K., Kitamura K. Flowering phenology, pollination, and fruit set of Cypripedium macranthos var. rebunense, a threatened Lady’s slipper (Orchidaceae) // Journal of Plant Research. – 2001. – Vol. 114, N 2. – P. 171–178.
Pijl L., Dodson С . H. Orchid Flowers: Their Pollination and Evolution. – Coral Gables: University of Miami Press, 1966. – 214 p.
Zhang Y., Zhao Sh, Liu D., Zhang Q., Cheng J. Flowering phenology and reproductive characteristics of Сypripedium macranthos (Orchidaceae) in China and their implication in conservation // Pakistan Journal of Botany. – 2014. – Vol. 46, N 4. – P. 1303–1308.
Svolynskiy A. D., Ivanov S. P., Kuramova V. V. Peculiarities of anthecology of Dactylorhiza romana (Orchidaceae) in Crimea: distribution, phenology, spatial distribution and morphometric of plants in flower // Ekosistemy. 2023. Iss. 33. P. 119–133.
The distribution of the orchid Dactylorhiza romana on the Crimean Peninsula has been analysed. On the southern coast of Crimea, the species is found throughout the entire western South Coast, and only at two sites in the east of the eastern South Coast (Karadag). In the Crimean Mountain forest zone D. romana is found only in the western parts of the southern and northern macroslopes of the Crimean Mountains, with the species rising to 1100 m.a.s.l. in the northern macroslope. In the Crimean foothill zone D. romana is found only at the border of the mountain forest zone in three separate localities in the western part of the foothills, one in the central part, and three in the far easternmost part of this natural zone. The phenodates of the flowering vary depending on the weather conditions of the season. The start of flowering is from 5 to 10 April and the end of flowering is from 10 to 20 May. Phenodates of flowering of South Coast and mountain-forest cenopopulations coincide, with the sum of positive temperatures of the beginning of flowering of cenopopulations on the South Coast of the Black Sea being 650 °С, and in mountain forests – 200 °С. The distribution of generative individuals in the two habitats in the Crimean Mountain forests (Osinovaya beam) and on the South Coast (Kastel mountain) is extremely irregular, from dense aggregations in which the distance between flowering individuals is 1–50 cm, to sparse with a distance between individuals 1–3 m or more. This distribution deviates strongly from the normal distribution and represents a MacArthur «broken stick» distribution. The South Coast and mountain-forest cenopopulations of D. romana differ significantly in the length of peduncles, inflorescences and the number of flowers in the inflorescences. Generative specimens from Mount Kastel are superior to those from Osinovaya beam in all of these parameters. Plants from the southern coastal locality are also characterized by higher values of the coefficient of variation in almost all parameters. Substantial differences in the sum of positive temperatures determining the phenodata of the flowering period, as well as morphological differences between plants of the South Coast and mountain-forest cenopopulations are interpreted as the presence in Crimea of two ecological forms of D. romana.
Key words: Dactylorhiza romana, Orchidaceae, distribution in Crimea, spatial distribution of individuals in the cenopopulation, flowering phenology, morphometric parameters of inflorescences, Crimean Peninsula.
Поступила в редакцию 23.02.23
Принята к печати 29.03.23
133
