
ОРНИТОГЕОГРАФИЧЕСКОЕ РАЙОНИРОВАНИЕ КРЫМСКОГО ПОЛУОСТРОВА
ORNITHOGEOGRAPHIC ZONING OF THE CRIMEAN PENINSULA
JOURNAL: «EKOSISTEMY», Issue 38, 2024
Publication text (PDF): Download
UDK: 598.2 (477.75)
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Kostin S. Yu.
Federal State Budgetary Institute of Science «The Order of the Red Banner of Labour Nikita Botanical Gardens — National Scientific Center RAS», Yalta, Russia
TYPE: Article
DOI: https://doi.org/10.29039/2413-1733-2024-38-90-100
PAGES: from 90 to 100
STATUS: Published
LANGUAGE: Russian
KEYWORDS: fauna, ornithocomplexes, geographical and genetic analysis, zoogeography, the Crimean Peninsula.
ABSTRACT (ENGLISH):
As a result of a comprehensive analysis of the avifauna of Crimea 21 districts were identified, taking into account the distribution of ecological and faunal groupings of birds according to zonal and biotopic divisions, the scheme of their dominance, the study of the relationships between species of various faunal complexes and types, as well as the presence of specific species. The border of the Sahara-Gobi and European subregions runs along the northern foothills. The species of Nomadic-type and tropical groups predominate in the lowland Crimea, and species of Nemoral complexes of European and Euro-Chinese types predominate in the mountain-forest belt. The northern foothills which are characterized by the transitional nature of the fauna that bears significant similarities with lowland Crimea (53 %), are attributed to the Crimean Foothills District. A belt of hemixerophytic forests, woodlands, tomillaries and savannoids of the Southern coast forms the Southern Coastal District. Both districts are considered part of the Western European Province of the European Subregion.
Введение
Географическое положение Крыма, находящегося в узле сопряжения нескольких природных зон, его сложное геологическое прошлое, ландшафтное и биологическое разнообразие обусловливают значительную гетерогенность авифауны полуострова, определяя формирование орнитокомплексов и эколого-фаунистических группировок, отличающихся по составу и происхождению.
Простое распределение числа видов и особей по типам фауны означает попытку осмыслить список видов в историческом аспекте, поскольку понятие о типах фауны птиц разработано с учетом их происхождения (Жуков, 2004). Отраслевое районирование (физико-географическое, ландшафтное, эдафо-климатическое, геоботаническое, зоогеографическое) природно-территориальных комплексов имеет большое практическое значение, так как оно лежит в основе ресурсоведения, природопользования и заповедного дела (Белик, 1996), что и определяет обширную библиографию, посвященную этому направлению исследований. При этом зоогеографическое районирование Крымского полуострова и, в частности, орнитогеографическое не разработано. Это определяет цель исследования – составить и охарактеризовать схему орнитогеографического районирования на основе региональных исторических связей.
МАТЕРИАЛ И МЕТОДЫ
В основу работы положены собственные материалы, собранные в 1980–2023 годах. Методологической основой работы является концепция «типов фаун» в трактовке Г. В. Никольского (1947), которая на орнитологическом материале получила развитие в работах Л. С. Степаняна (1967), А. А. Назаренко (1968), А. А. Кищинского (1977, 1980, 1988), В. П. Белика (1996, 2006, 2013) и других орнитологов. Поскольку основным критерием отнесения вида птиц к одному из типов фауны является конфигурация гнездового ареала (Штегман, 1938), в настоящей работе мы анализируем гнездовую фауну птиц полуострова.
Определяющее значение при делении внеарктической части Палеарктики имеют подходы к районированию: на основе зональных принципов или региональных исторических связей. В настоящее время практически все схемы деления Палеарктики на зоогеографические подобласти основаны на региональном принципе, где выделяют 4 подобласти: Сибирскую, Европейскую, Китайскую, а также Сахаро-Гобийскую. Соответственно этим подобластям выделяют 4 типа фауны: Сибирский, Европейский, Китайский и Номадийский. Южные горы целесообразно выделять в особую подобласть с характерным для нее типом фауны, к которому следует относить лишь обитателей субальпики (криволесий и кустарников) и альпийской зоны (лугов и скал), составляющих два отдельных больших фаунистических комплекса (Белик, 2006, 2013).
В последние полвека в зоогеографических работах (Назаренко, 1968; Кищинский, 1977, 1980, 1988; Белик, 1996) было показано, что методы анализа на основе типов фауны пригодны для изучения зооценозов крупных хоронов – областей, подобластей, тогда как региональные фауны продуктивно анализировать, используя категорию фаунистических комплексов, связанных происхождением не с подобластями, а ландшафтными зонами. Биоценотическая специфика фаунистических комплексов проявляется посредством выявления эколого-фаунистических группировок, адаптированных к определенным биотопам в различных типах ландшафта (табл. 1).
Таблица 1
Типы фаун, фаунистические комплексы и эколого-фаунистические группировки Палеарктики (по Белику, 2006 с дополнениями)
|
Типы фаун |
Фаунистические комплексы |
Эколого-фаунистические группировки |
Интразональные эколого-фаунистические группировки |
|
Европейский |
Неморальный |
Собственно неморальная |
Аллювиофильно-луговая |
|
Болотно-неморальная |
|||
|
Борово-неморальная |
|||
|
Лесостепной |
Собственно лесостепная |
||
|
Пойменно-лесолуговая |
|||
|
Горно-лесостепная |
|||
|
Субсредиземно-морский |
Ксерофильно-дубравная |
||
|
Горно-хвойная |
|||
|
Ксерофильно-кустарниковая |
|||
|
Евро-Китайский |
Древне-неморальный |
Собственно древне-неморальная |
|
|
Болотно-древне-неморальная |
|||
|
Борово-древне-неморальная |
|||
|
Древне-лесостепной |
Собственно древне-лесостепная |
||
|
Пойменно-древне-лесолуговая |
|||
|
Горно-древне-лесостепная |
|||
|
Номадийский |
Пустынно-степной |
Пустынно-петрофитная |
|
|
Сухо-степная |
|||
|
Пустынно-горный |
Склерофильная |
||
|
Лиманный |
Аллювиофильно-галофитная |
||
|
Лиманно-островная |
|||
|
Плавневая |
В связи с этим результаты анализа зонально-биотопического распределения эколого-фаунистических группировок Крыма (Ю. Костин и др., 1999; Бескаравайный, 2001; Костин, 2014) составляют одну из основ схемы орнитогеографического районирования региона. Элементами в предлагаемой нами схеме зоогеографического анализа (рис. 1) кроме биотопов (местообитаний) выступают региональные соотношения типов фаун, фаунистических комплексов и эколого-фаунистических группировок. При этом основным методом орнитогеографического анализа остается сравнение гнездовой фауны отдельных биотопических выделов Крыма, как между собой, так и с фауной смежных и отдаленных регионов, а также сопоставление их географо-генетической структуры с применением коэффициента Жаккара и кластерного анализа (Костин, 2023).
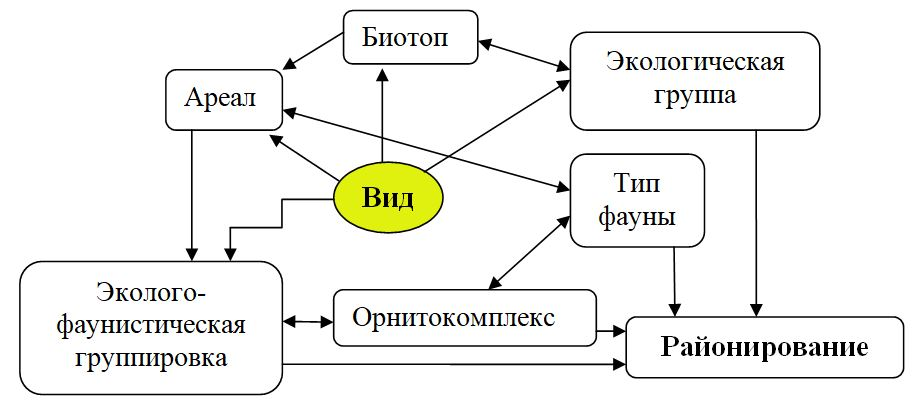
Рис. 1. Схема зоогеографического анализа авифауны Крыма
Систематический порядок и таксономия птиц приведены в соответствии со списком Е. А. Коблика, В. Ю. Архипова (2014).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Крым находится на западной окраине суббореальных степей умеренного пояса, где зональным является семиаридный степной тип ландшафта, характерный для равнинной части полуострова. В предгорьях и горах формируются другие типы ландшафтов, что связано с барьерно-высотной поясностью и позиционностью относительно вещественно-энергетических потоков и, как следствие, другие по сравнению с равнинами гидротермический режим, эдафо-климатические и фитоценотические характеристики (Современные ландшафты…, 2009). Ландшафтное и биотопическое разнообразие полуострова определяет существование зональных, азональных и интразональных комплексов, мозаичное пространственное распределение значительного количества эколого-фаунистических группировок.
В основу районирования равнинного Крыма положены особенности соотношения между представителями Номадийского, Европейского типов фауны и тропической группы, относящихся к пустынно-степному, пустынно-горному фаунистическим комплексам и интразональным эколого-фаунистическим группировкам, с учетом участия дендрофилов неморального и лесостепного комплексов. В равнинном Крыму нами выделено 10 районов (рис. 2), которые можно разделить на две группы. К первой группе относятся: Каркинитский, Центральный и Восточный Присивашские, Донузлавско-Межводненский, Сакско-Евпаторийский районы. По зонально-биотопическому делению все они расположены в гидроморфном ландшафтном уровне в подзоне полупустынных степей (табл. 2, рис. 3). Районы различаются степенью участия видов интразональных эколого-фаунистических группировок: плавневой, лиманно-островной и аллювиофильно-галофитной. В этой группе доминантами являются лимнофилы, относящиеся к Номадийскому типу фауны и тропической группе.
В Каркинитский район (Ст-1) входит территория от Бакальской косы до Красноперекопска, но большая часть соленых озер западного Сиваша занята отстойниками химического производства. На остальной территории побережья Каркинитского залива, Лебяжьих о-вах находятся наиболее стабильное поселение (≈40 пар) длинноносого крохаля Mergus serrator Linnaeus, 1758 (северо-таежный комплекс Сибирского типа); постоянные колонии голенастых и чайковых, в частности чегравы Hydroprogne caspia (Pallas, 1770), черноголового хохотуна Larus ichthyaetus Pallas, 1773, в 2000–2002 годах гнездился розовый пеликан Pelеcanus onocrotalus Linnaeus, 1758. В береговой полосе Кумовских плавней гнездится малый баклан Phalacrocorax pygmeus (Pallas, 1773); отмечена наибольшая концентрация белощекой крачки Chlidonias hybrida (Pallas, 1811), белого аиста Ciconia ciconia (Linnaeus, 1758) и черногрудого воробья Passer hispaniolensis (Temminck, 1820). У Красноперекопска в колонии грача Corvus frugilegus Linnaeus, 1758 зарегистрировано гнездование египетской цапли Bubulcus ibis (Linnaeus, 1758), на останцевых островах Айгульского и Кирлеутского озер – красавки Anthropoides virgo (Linnaeus, 1758), на Бакальской косе – большого кроншнепа Numenius arquata (Linnaeus, 1758), а в тростниках Айгульского озера – единственное в Крыму место гнездования речного сверчка Locustella fluviatilis (Wolf, 1810).
В Присивашских районах широко представлены островные системы и галофитные луга, в которых формируется лиманно-островная и аллювиофильно-галофитная группировки, у сбросов ирригационной сети Северо-Крымского канала в Сиваш, где были развиты обширные тростниково-рогозовые заросли, распространены виды плавневой группировки. Вдоль транспортных магистралей в лесомелиоративных массивах гнездятся в основном представители собственно лесостепной и пойменно-лесолуговой группировок Европейского типа фауны.
Центральный Присивашский район отличается преобладанием видов сухо-степной группировки пустынно-степного и лиманного комплексов при доминировании представителей аллювиофильно-галофитной группировки с наличием специфичного вида – серого жаворонка Calandrella rufescens (Vieillot, 1819), а у о-ва Чурюк в 2017 году отмечено гнездование фламинго Phoenicopterus roseus Pallas, 181. Но при этом только в этом районе на останцевых островах отмечено спорадическое гнездование широконоски Anas clypeata Linnaeus, 1758 и шилохвости Anas acuta Linnaeus, 1758 – видов гипоарктического комплекса. Здесь располагаются основные поселения шилоклювки Recurvirostra avosetta Linnaeus, 1758, морского голубка Larus genei Breme, 1840, пестроносой крачки Thalasseus sandvicensis (Latham, 1787). В отличие от других районов северного и северо-восточного Крыма здесь в береговой полосе широко представлены глинистые обрывы (до 7–10 м), где склерофильную группировку составляют 11 видов.
Восточный Присивашский район (Ст-3) характеризуется многочисленными аккумулятивными островами, на которых формируется лиманно-островная группировка. Здесь расположены крупные балочные комплексы в устьевых зонах, занятые обширными тростниковыми крепями, в которых выявлены наиболее многочисленные поливидовые колонии плавневой группировки; находятся основные крымские поселения малого баклана, желтой Ardeola ralloides (Scopoli, 1769) и рыжей Ardea purpurea Linnaeus, 1766 цапель, кваквы Nycticorax nycticorax (Linnaeus, 1758), каравайки Plegadis falcinellus (Linnaeus, 1766), серого гуся Anser anser (Linnaeus, 1758), красноносого нырка Netta rufina (Pallas, 1773). На территории Калиновского природного парка, занимающего урочище Калиновка между полуостровами Тюп-Тархан и Стефановский, представлены виды сухо-степной группировки, а вдоль канала в лесомелиоративных посадках – виды лесостепного комплекса.
Донузлавско-Межводненский район (Ст-5) выделяется сочетанием видов интразональных эколого-фаунистических группировок с доминированием представителей Номадийского типа фауны: аллювиофильно-галофитной, плавневой, склерофильной; с незначительным участием видов аллювиофильно-луговой и дендрофилов собственно лесостепной группировок, включающей представителей Европейского типа фауны. Район делится на два участка, отличающихся особенностями мезорельефа. Большую часть Донузлавского участка занимает акватория одноименного озера (глубиной до 27 м), берега
Таблица 2 (на следующей странице)
Фауно-генетическая структура орнитокомплексов отдельных орнитогеографических районов Крыма
|
Районы |
Типы фаун |
Тропические группы |
Всего видов |
||||||||||||
|
Европейский |
Евро-Китайский |
Номадийский |
Сибирский |
Гималайский |
|||||||||||
|
Фаунистические комплексы (количество видов) Орнито-географические районы |
Неморальный |
Лесостепной |
Суб-средиземно-морский |
Аллювио-фильный |
Древне-неморальный |
Древне-лесостепной |
Пустынно-степной |
Пустынно-горный |
Лиманный |
Горно-таежный |
Северо-таежный |
Суб-альпийский |
|||
|
Степные |
Каркинитский |
5 |
9 |
1 |
9 |
1 |
4 |
9 |
11 |
27 |
0 |
1 |
0 |
31 |
108 |
|
Центральный Присивашский |
4 |
7 |
1 |
4 |
1 |
2 |
7 |
11 |
21 |
0 |
1 |
0 |
21 |
86 |
|
|
Восточный Присивашский |
4 |
8 |
1 |
9 |
1 |
3 |
9 |
9 |
29 |
0 |
0 |
0 |
29 |
103 |
|
|
Тарханкутский |
3 |
10 |
3 |
1 |
0 |
3 |
8 |
15 |
4 |
0 |
0 |
0 |
9 |
59 |
|
|
Донузлавско-Межводненский |
3 |
7 |
3 |
4 |
1 |
2 |
6 |
14 |
10 |
0 |
0 |
0 |
16 |
67 |
|
|
Сакско-Евпаторийский |
7 |
12 |
2 |
8 |
1 |
4 |
8 |
15 |
20 |
0 |
0 |
0 |
17 |
95 |
|
|
Центрально-степной |
7 |
11 |
2 |
1 |
0 |
5 |
6 |
12 |
3 |
0 |
0 |
0 |
5 |
52 |
|
|
Индольский |
8 |
12 |
2 |
7 |
1 |
4 |
5 |
12 |
12 |
0 |
0 |
0 |
11 |
75 |
|
|
Керченский приазовский |
7 |
13 |
4 |
8 |
1 |
5 |
10 |
18 |
21 |
0 |
0 |
0 |
22 |
110 |
|
|
Керченский при-черноморский |
8 |
13 |
3 |
23 |
2 |
3 |
9 |
17 |
8 |
0 |
0 |
0 |
17 |
86 |
|
|
Предгорные |
Гераклейский |
9 |
14 |
4 |
2 |
2 |
3 |
6 |
16 |
4 |
0 |
0 |
0 |
8 |
71 |
|
Бахчисарайско-Белогорский |
12 |
13 |
4 |
4 |
7 |
3 |
3 |
14 |
4 |
1 |
0 |
0 |
9 |
75 |
|
|
Северо-Восточный предгорный |
12 |
16 |
4 |
3 |
7 |
6 |
5 |
14 |
4 |
0 |
0 |
0 |
14 |
86 |
|
|
Горно-лесные |
Северный горно-лесной |
11 |
5 |
3 |
3 |
10 |
3 |
0 |
4 |
0 |
2 |
1 |
1 |
4 |
48 |
|
Юго-Западный горно-лесной |
14 |
6 |
1 |
3 |
6 |
1 |
0 |
3 |
0 |
3 |
1 |
0 |
4 |
43 |
|
|
Юго-восточный горно-лесной |
12 |
4 |
0 |
3 |
7 |
1 |
0 |
4 |
0 |
1 |
0 |
0 |
4 |
37 |
|
|
Западный нагорный |
8 |
5 |
0 |
1 |
1 |
2 |
1 |
8 |
0 |
2 |
0 |
0 |
3 |
32 |
|
|
Восточный нагорный |
7 |
6 |
0 |
3 |
1 |
4 |
3 |
7 |
0 |
1 |
0 |
1 |
3 |
37 |
|
|
Суб-средиземно-морские |
Западный южно-бережный |
8 |
8 |
2 |
2 |
3 |
1 |
0 |
8 |
2 |
0 |
0 |
0 |
5 |
41 |
|
Центральный южно-бережный |
8 |
11 |
3 |
2 |
4 |
2 |
1 |
10 |
3 |
0 |
0 |
0 |
9 |
54 |
|
|
Юго-Восточный |
9 |
14 |
4 |
5 |
8 |
2 |
6 |
16 |
14 |
0 |
0 |
0 |
14 |
95 |
|
которого в приморской части образуют ракушечно-песчаную косу, в центральной части изобилуют скальные выходы, а верховья покрыты обширными тростниковыми зарослями. К особенностям населения птиц этого участка можно отнести локальное поселение плешанки Oenanthe pleschanka (Lepechin, 1770) и существование до середины 1970-х годов колоний степной пустельги Falco naumanni Fleischer, 1818 по скалистым берегам озера. Поселения голенастых в пресноводных верховьях появились в последние десятилетия и отличаются низкими показателями видового разнообразия (большая белая Casmerodius alba (Linnaeus, 1758) и серая Ardea cinerea Linnaeus, 1758 цапли) и численности – 6–14 пар. При относительно полном видовом составе плавневой группировки (поганки, гусеобразные, камышевки) отсутствует красноносый нырок. Межводненский участок находится на северном побережье Тарханкута и кроме береговых обрывов с поселениями хохлатого баклана Phalacrocorax aristotelis (Linnaeus, 1761), балобана Falco cherrug J. E. Gray, 1834, хохотуньи Larus cachinnans Pallas, 1811, включает горько-соленые озера лиманного типа (Ярылгач, Панское) с видами аллювиофильно-галофитной группировки. В верховьях оз. Джарылгач сформировались обширные пресноводные мелководья, заселенные представителями аллювиофильно-луговой группировки.
Сакско-Евпаторийский район отличается преобладанием урбо-селитебных и промышленных комплексов с синантропной фауной в сочетании с представителями плавневой группировки – лебедь-шипун Cygnus olor (J.F. Gmelin, 1789), кряква Anas
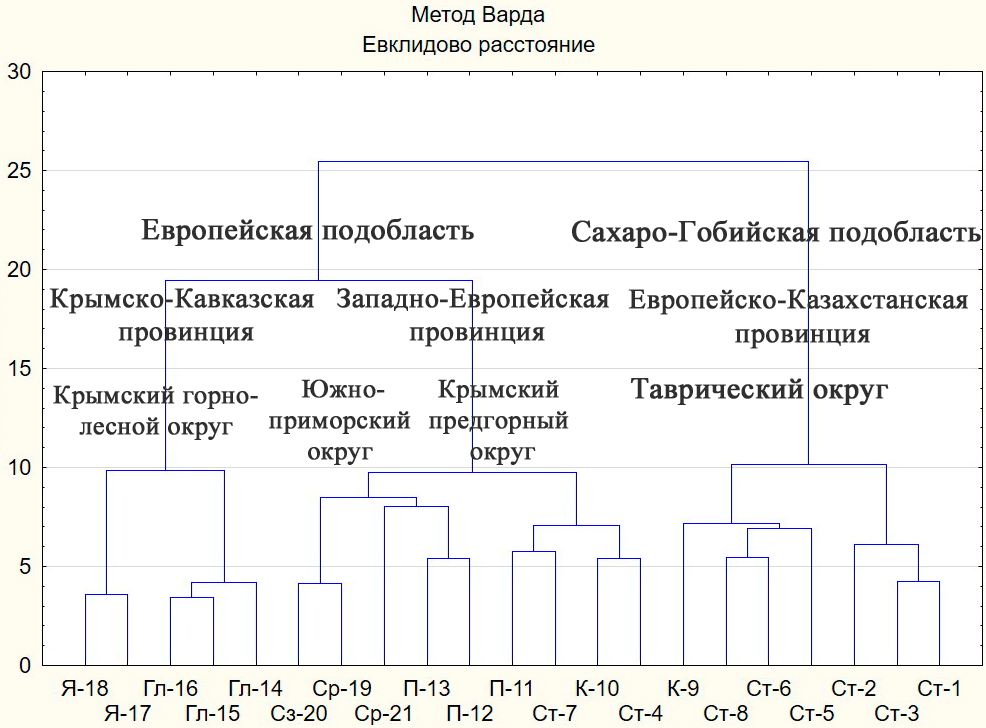
Рис. 2. Результаты кластерного анализа по выделению орнитофаунистических районов
Равнинный Крым: I. Степные районы: 1. Каркинитский (Ст-1). 2. Центральный Присивашский (Ст‑2). 3. Восточный Присивашский (Ст-3). 4. Тарханкутский (Ст-4). 5. Донузлавско-Межводненский (Ст-5). 6. Сакско-Евпаторийский (Ст-6). 7. Центрально-степной (Ст-7). 8. Индольский (Ст-8). II. Керченские районы: 9. Приазовский (К-9). 10. Причерноморский (К-10).
Горный Крым: III. Предгорные районы: 11. Гераклейский (П-11). 12. Бахчисарайско-Белогорский (П-12). 13. Северо-восточный (П-13). IV. Горно-лесные районы: 14. Северный (Гл-14). 15. Юго-западный (Гл-15). 16. Юго-восточный (Гл-16). V. Нагорные районы (яйлы): 17. Западный (Я-17). 18. Восточный (Я-18). VI. Субсредиземноморские районы: 19. Западный (Ср-19). 20. Центральный (Ср‑20). 21. Юго-восточный (Ср-21).
platyrhynchos Linnaeus, 1758, чирок-трескунок Anas querquedula Linnaeus, 1758, красноголовый Aythya ferina (Linnaeus, 1758) и белоглазый A. nyroca (Güldenstädt, 1770) нырки. Специфическим видом района выступает савка Oxyura leucocephala (Scopoli, 1769).
Во вторую группу входят Тарханкутский; Центрально-степной; Индольский; Керченские Приазовский и Причерноморский районы. Все они находятся в плакорном ландшафтном уровне, где преобладают участки, занятые полями и пастбищами. Настоящие ковыльные и разнотравные степи представлены фрагментарно.
В Центрально-степном районе (Ст-7) значительные площади заняты агроценозами, развитыми селитебными и урбо-промышленными биотопическими комплексами. Он характеризуется отсутствием стенотопных кампофилов (дрофа Otis tarda Linnaeus, 1758, стрепет Tetrax tetrax (Linnaeus, 1758), красавка) и минимальным участием склерофилов пустынно-горного комплекса. Здесь проходит южная граница регионального ареала соловья Luscinia luscinia (Linnaeus, 1758), а в северо-восточной части района находится локальная гнездовая группировка орла-могильника Aquila heliaca Savigny, 1809.
Выделение Керченских районов обусловлено присутствием стрепета, черноголовой овсянки Granativora melanocephala (Scopoli, 1769), осоеда Pernis apivorus (Linnaeus, 1758), степной пустельги, крупной группировки черноголового чекана Saxicola rubicola (Linnaeus, 1766). Приазовский район (К-9) отличается более мезофильными условиями и значительными
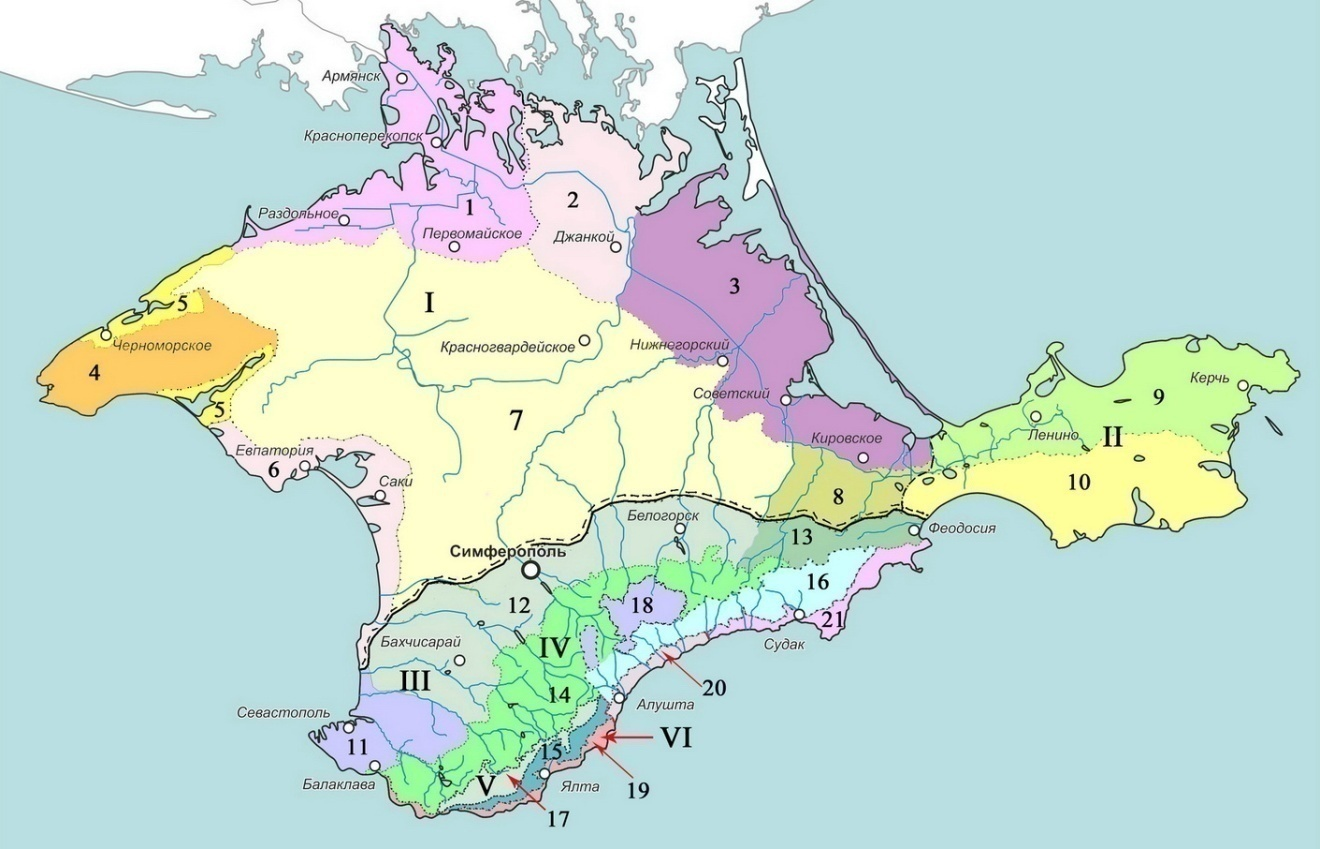
Рис. 3. Схема орнитогеографического районирования Крыма
Равнинный Крым: I. Степные районы: 1. Каркинитский (Ст-1). 2. Центральный Присивашский (Ст‑2). 3. Восточный Присивашский (Ст-3). 4. Тарханкутский (Ст-4). 5. Донузлавско-Межводненский (Ст-5). 6. Сакско-Евпаторийский (Ст-6). 7. Центрально-степной (Ст-7). 8. Индольский (Ст-8). II. Керченские районы: 9. Приазовский (К-9). 10. Причерноморский (К-10).
Горный Крым: III. Предгорные районы: 11. Гераклейский (П-11). 12. Бахчисарайско-Белогорский (П-12). 13. Северо-восточный (П-13). IV. Горно-лесные районы: 14. Северный (Гл-14). 15. Юго-западный (Гл-15). 16. Юго-восточный (Гл-16). V. Нагорные районы (яйлы): 17. Западный (Я-17). 18. Восточный (Я-18). VI. Субсредиземноморские районы: 19. Западный (Ср-19). 20. Центральный (Ср‑20). 21. Юго-восточный (Ср-21).
по площади плавнями (лебедь-шипун, серый гусь), тогда как в восточной части района (Караларская степь) сохранился полный состав пустынно-степного комплекса (стрепет, дрофа, красавка, авдотка Burhinus oedicnemus (Linnaeus, 1758)). Причерноморский район (К‑10) характеризуется присутствием средиземноморской чайки Larus michahellis J. F. Naumann, 1840, черноухой каменки Oenanthe melanoleuca (Güldenstädt, 1775), черногрудого воробья и колоний хохлатого баклана. Тарханкутский район (Ст-4) выделяется сочетанием скально-морских (хохлатый баклан, сапсан Falco peregrinus Tunstall, 1771) и пустынно-степных (дрофа, красавка) биотопов. В связи с наличием приморских оползневых «цирков», береговых обрывов и развитой овражно-балочной системы в Тарханкутском (Ст-4) и Керченском причерноморском (К-10) районах представлено большинство видов склерофильной группировки, что определяет их объединение в одну ветвь дендрограммы (рис. 2).
Индольский район (Ст-8) отличается смешанной фауной, специфику которой определяет его положение между северными предгорьями, Присивашьем и Центрально-степным районом. Поэтому в составе его фауны сочетаются виды неморального и лесостепного комплексов (осоед, могильник), ксерофильно-кустарниковой (южный соловей Luscinia megarhynchos C.L.Brehm, 1831) и ксерофильно дубравной (сирийский дятел Dendrocopos syriacus (Hemprich et Ehrenberg, 1833)) группировок с видами сухо-степной и плавневой группировок.
Семь районов по преобладанию представителей Номадийского типа фауны, формирующие разные эколого-фаунистические группировки пустынно-степного и лиманного фаунистических комплексов, рассматриваются нами в составе Таврического округа Европейско-Казахстанской провинции Сахаро-Гобийской подобласти. Отнесение большинства районов равнинного Крыма к данной провинции подтверждается значительным сходством по коэффициенту Жаккара фаун равнинного Крыма в целом с фаунами Калмыкии (66 %), Степного Подонья (65 %), предкавказского Ставрополья (62 %) и Гурьевской области Казахстана (58 %). По нашему мнению, к Таврическому округу вместе с Таманским районом, выделенного В. П. Беликом (2013), относятся Приазовские лиманы, Присивашье и северо-восточное Причерноморье, где наиболее выражены фаунистические комплексы морских островов, солончаков, песчаных кос. Керченский причерноморский, Тарханкутский и Центрально-степной районы по видовому составу и эколого-фаунистическим группировкам отличаются от других районов, выделенных в равнинном Крыму, так как в них наиболее выражены склерофильная и сухо-степная группировки.
В основу районирования горного Крыма положены особенности соотношения между склерофилами пустынно-горного и дендрофилами лесостепного, неморального и средиземноморского фаунистических комплексов, относящихся к Европейскому типу фауны. В результате комплексного анализа в горном Крыму выделены 11 районов (рис. 2).
Пояс предгорий, в котором выделено 3 района, характеризуется мозаичностью биотопической структуры и преимущественно населен лесостепными дендрофилами и склерофилами. Гераклейский район (П-11) отличается преобладанием представителей селитебно-промышленных биотопов (домовый сыч Athene noctua (Scopoli, 1769), сорока Pica pica (Linnaeus, 1758), галка Corvus monedula Linnaeus, 1758) с участием видов, характерных для околоводных (пеганка Tadorna tadorna (Linnaeus, 1758), красноголовый нырок Aythya ferina (Linnaeus, 1758)), скально-морских (хохлатый баклан), скальных (белоголовый сип Gyps fulvus (Hablizl, 1783), сапсан), борово-неморальных (большой пестрый дятел Dendrocopos major (Linnaeus, 1758), пеночка-трещетка Phylloscopus sibilatrix (Bechstein, 1793), деряба Turdus viscivorus Linnaeus, 1758) и горно-лесостепных (вяхирь Columba palumbus Linnaeus, 1758, горлица Streptopelia turtur (Linnaeus, 1758), щегол Carduelis carduelis (Linnaeus, 1758), коноплянка Acanthis cannabina (Linnaeus, 1758)) местообитаний.
Бахчисарайско-Белогорский район (П-12) занимает большую часть северных предгорий, для него характерно сочетание видов сухо-степной (перепел Coturnix coturnix (Linnaeus, 1758), красавка); собственно лесостепной (серая куропатка Perdix perdix (Linnaeus, 1758), чеглок Falco subbuteo Linnaeus, 1758, могильник, осоед, вертишейка Jynx torquilla Linnaeus, 1758, иволга Oriolus oriolus (Linnaeus, 1758), сорока, грач), склерофильной (курганник Buteo rufinus (Cretzschmar, 1827), галка, золотистая щурка Merops apiaster Linnaeus, 1758, белобрюхий Apus melba (Linnaeus, 1758) и черный A. apus (Linnaeus, 1758) стрижи), а также плавневой (огарь Tadorna ferruginea (Pallas, 1764), кряква, лысуха Fulica atra Linnaeus, 1758, ремез Remis pandelinus (Linnaeus, 1758), усатая синица Panurus biarmicus (Linnaeus, 1758)) группировок. Специфическим видом района является зимородок Alcedo atthis (Linnaeus, 1758).
Северо-восточный район (П-13) отличается большей лесистостью (змееяд Circaetus gallicus (J.F. Gmelin, 1788), вальдшнеп Scolopax rusticola Linnaeus, 1758) в сочетании с лесостепью (грач, южный соловей, обыкновенная Emberiza citrinella Linnaeus, 1758, садовая E. hortulana Linnaeus, 1758 овсянки) в западной части и безлесным холмогорьем с полями зерновых и виноградниками на востоке.
Предгорья Крыма вместе с Новороссийским районом Черноморского побережья Кавказа В. П. Белик (2013) предложил отнести к Эвксинскому округу в составе Сумерийской провинции по значительному участию пустынно-горных видов в сложении фаунистических комплексов. По нашим данным, фауна крымских предгорий является преимущественно лесостепной, занимая вторичные лесо-кустарниковые биотопы, сады и виноградники. Поэтому она отвечает признакам Лесостепной переходной зоны, и описанные районы отнесены нами не к Сумерийской провинции, а к Крымскому предгорному округу Западно-Европейской провинции Европейской подобласти.
В эту же провинцию и подобласть, но в Южно-приморский округ, отнесены три субсредиземноморских района. Западный район (Ср-19) отличается сочетанием селитебных ландшафтов, парково-рекреационных комплексов с преобладанием интродуцентов (кипарисы, кедры, платан и др.) и гемиксерофитных редколесий, кустарниковых сообществ. Специфическим видом здесь является красноголовый королек Regulus ignicapillus (Temminck, 1820), а фоновыми – синантропные виды пустынно-горной (стрижи, ласточки, воробьи) и лесостепной (горлица Streptopelia turtur (Linnaeus, 1758), сплюшка Otus scops (Linnaeus, 1758), ворона Corvus corax Linnaeus, 1758, ополовник Aegithalos caudatus (Linnaeus, 1758), зеленушка Chloris chloris (Linnaeus, 1758), щегол Carduelis carduelis (Linnaeus, 1758)) группировок с участием лесных (сойка Garrulus glandarius (Linnaeus, 1758), серая неясыть Strix aluco Linnaeus, 1758), субсредиземноморских (хохлатый баклан, средиземноморская чайка, серая славка Sylvia communis Latham, 1787) видов (Костин, 2014, 2021).
К востоку от Алуштинской долины до Судака в приморской полосе расположен Центральный район (Ср-20), характеризующийся более ксерофитными условиями, преобладанием кустарникового типа растительности на каменистых склонах, отсутствием крупных рекреационно-парковых комплексов. В составе фауны отсутствуют хохлатый баклан, хохотунья Larus cachinnans Pallas, 1811 и средиземноморская чайка, появляются малый зуёк Charadrius dubius Scopoli, 1786, удод Upupa epops Linnaeus, 1758, ястребиная славка Sylvia nisoria (Bechstein, 1792), а также отсутствующие в Западном районе домовый сыч, сорока.
Биотическая неоднородность Юго-Восточного района (Ср-21) определяет наличие ряда специфических черт фаунистических комплексов. Появление лимнофилов (поганки, кряква, серый гусь, лысуха, пеганка, огарь, дроздовидная камышевка Acrocephalus arundinaceus (Linnaeus, 1758) и др.) обусловлено существованием здесь искусственных пресных водоемов. С аридно-петрофитными биотопами полуострова Меганом связаны каменка-плясунья, с грунтовыми обнажениями – золотистая щурка и сизоворонка Coracias garrulus Linnaeus, 1758, скальными обнажениями – хохлатый баклан, хохотунья и средиземноморская чайка (Бескаравайный, 2001).
Значительное участие видов Европейского типа фауны с небольшим количеством субсредиземноморских видов (7) в приморской полосе гемиксерофитных лесов, редколесий, томилляров и саванноидов определило выделение нами Южно-приморского округа и отнесение его к Западно-Европейской, а не в Субсредиземноморской провинции, характеризующейся по Н. Н. Щербаку (1988) значительным участием средиземноморских элементов фауны беспозвоночных и рептилий.
Горно-лесные районы расположены в среднегорном уровне в поясе высокоствольных дубовых, буковых и сосновых лесов. Их различия определяются поясностью, эдафо-климатическими условиями, составом растений-эдификаторов. Северный район (Гл-14) занимает северный макросклон Главной гряды с более гумидными условиями и развитыми высокоствольными дубовыми и буковыми лесами, где ядро авифауны составляют виды собственно неморальной и борово-неморальной группировок, а специфичными видами являются черный аист Ciconia nigra (Linnaeus, 1758), черный гриф Aegypius monachus (Linnaeus, 1766), вальдшнеп, клинтух Columba oenas Linnaeus, 1758, полуошейниковая мухоловка Ficedula semitorquata (Homeyer, 1885), пеночка-теньковка Phylloscopus collybita (Vieillot, 1817). Юго-западный район (Гл-15) отличается присутствием видов горно-таежного комплекса (московка Parus ater Linnaeus, 1758, чиж Spinus spinus (Linnaeus, 1758), клест Loxia curvirostra Linnaeus, 1758) и доминированием видов борово-неморальной группировки. В Юго-восточном районе (Гл-16) более представлены ксерофитные местообитания, для которых характерны лесной жаворонок Lullula arborea (Linnaeus, 1758), поэтому меньше участие стенотопных сильвантов, к специфичным видам относится красноголовый сорокопут Lanius senator Linnaeus, 1758) (Бескаравайный, 2001; Костин, 2014, 2021).
Специфичным видом нагорных районов (яйл) является пестрый каменный дрозд Monticola saxatilis (Linnaeus, 1766), но по численности здесь доминируют жаворонки, а в полосе прияйлинских криволесий – виды горно-лесостепной группировки: лесной конек Anthus trivialis (Linnaeus, 1758), коноплянка, зеленушка Chloris chloris (Linnaeus, 1758), черноголовая славка Sylvia atricapilla (Linnaeus, 1758). Западные яйлы (Я-17) отличаются присутствием клеста, лесной завирушки Prunella modularis (Linnaeus, 1758), Восточные (Я‑18) – серой куропатки, кряквы, коростеля Crex crex (Linnaeus, 1758), удода в связи с развитыми каровыми полями (Ю. Костин и др., 1999).
Горно-лесные и нагорные районы Крыма, заселенные в основном дендрофилами Европейского и Евро-Китайского типов, а также широко распространенными синантропными склерофилами Номадийского типа фаун, мы относим в Крымский горно-лесной округ, выделяемый Б.А. Кузнецовым (1950), однако рассматриваем его в Крымско-Кавказской провинции Европейской подобласти.
ЗАКЛЮЧЕНИЕ
В результате комплексного анализа авифауны Крыма, с учетом распределения эколого-фаунистических группировок птиц по зонально-биотопическим выделам, соотношения между представителями различных фаунистических комплексов и типов фаун, а также наличия специфических видов выделен 21 район в составе 4 округов, 3 провинций и 2 подобластей. По северным предгорьям проходит граница крупных зоохоронов – Сахаро-Гобийской и Европейской подобластей. На это указывает низкий коэффициент сходства (22 %) гнездовых фаун равнинной и горно-лесной частей. В равнинном Крыму преобладают виды Номадийского типа и тропической группы, в горно-лесном поясе – виды неморальных комплексов Европейского и Евро-Китайского типов фаун. По видовому составу фауна переходного предгорного пояса имеет наибольшее сходство с фауной равнинного Крыма (53 %), тогда как с фауной среднегорного пояса сходство составляет 42 %. Отличия между фаунами предгорного и равнинного Крыма нивелирует антропогенное воздействие, связанное с лесо- и гидромелиоративной деятельностью в равнинном Крыму (Костин, 2023).
Работа выполнена в рамках выполнения тем государственного задания ФГБУН Никитского ботанического сада – Национального научного центра РАН.
Белик В. П. Орнитогеографическое районирование Степного Подонья: современная статика и динамика территориального распределения птиц // Беркут. – 1996. – Т. 5, вып. 2. – С. 111–124.
Белик В. П. Фауногенетическая структура авифауны Палеарктики // Зоологический журнал. – 2006. – Т. 85, № 3. – С. 298–316. DOI: 101134/S0013873806100022
Белик В. П. Орнитогеографические связи и районирование Большого Кавказа (новые подходы в анализе фауны) // Стрепет. – 2013. – Т. 1, № 1. – С. 5–88.
Бескаравайный М. М. Биотопическое распределение птиц восточной части Горного Крыма // Бранта: Сборник научных трудов Азово-Черноморской орнитологической станции. – 2001. – Вып. 4. – С. 42–70.
Жуков В. С. Хорологический анализ орнитофауны Северной Евразии: ландшафтно-экологический аспект. Аналитический обзор // Серия Экология. – Новосибирск, 2004. – Вып. 74. – 182 с.
Кищинский А. А. Принципы реконструкции истории авифаун биогеографическим методом // Адаптивные особенности и эволюция птиц. – М.: Наука, 1977. – С. 33–39.
Кищинский А. А. Птицы Корякского нагорья. – М.: Наука, 1980. – 336 с.
Кищинский А. А. Орнитофауна северо-востока Азии: История и современное состояние. – М.: Наука, 1988. – 288 с.
Коблик Е. А., Архипов В. Ю. Фауна птиц стран Северной Евразии в границах бывшего СССР: списки видов. Зоологические исследования, № 14. – М.: Товарищество научных изданий КМК, 2014. – 171 с.
Костин С. Ю. Каталог птиц Крыма. – Симферополь: ИТ АРИАЛ, 2020. – 244 с.
Костин С. Ю. Птицы Крымского природного заповедника // Научные записки природного заповедника «Мыс Мартьян». – 2014. – Вып. 5. – С. 122–204.
Костин С. Ю. Современная структура авифауны и особенности распределения птиц Крыма // Зоологический журнал. – 2021. – Т. 100, № 8. – С 897-913. DOI: 10.31857/S0044513421060088
Костин С. Ю. Особенности географо-генетической структуры фауны птиц Крыма // Бюллетень Государственного Никитского ботанического сада. – 2023. – Вып. 147. – С. 127–134. DOI: 10.25684/0513-1634-2023-147-127-134
Костин Ю. В., Дулицкий А. И., Костин С. Ю. Эколого-географическая характеристика зонально-биотопических выделов и состав их фауны // Вопросы развития Крыма: Научно-практический дискуссионно-аналитический сборник. Вып. 11: Биологическое и ландшафтное разнообразие Крыма: проблемы и перспективы. – Симферополь: Сонат, 1999. – С. 35–54.
Кузнецов Б. А. Очерк зоогеографического районирования СССР. – М.: Изд-во МОИП, 1950. – 176 с.
Назаренко А. А. Ценотические реликты и ландшафтная приуроченность неморальной орнитофауны юга Дальнего Востока // Орнитология. – 1968. – Вып. 9. – C. 121–130.
Никольский Г. В. О биологической специфике фаунистических комплексов и значении ее анализа для зоогеографии // Зоологический журнал. – 1947. – Т. 26, вып. 3. – С. 221–230.
Современные ландшафты Крыма и сопредельных акваторий / [Науч. ред. Е. А. Позаченюк]. – Симферополь: Бизнес-Информ, 2009. – 672 с.
Степанян Л. С. Горно-пустынная авифаунистическая группировка Передней Азии и ее орнитогеографическая оценка // Орнитология. – 1967. – Вып. 8. – C. 123–132.
Штегман Б. К. Основы орнитогеографического деления Палеарктики // Фауна СССР. Птицы. Т. 1, вып. 2. – М.-Л.: Изд-во АН СССР, 1938. – 156 с.
Щербак Н. Н. Зоогеографическое деление Украинской ССР // Вестник зоологии. – 1988. – № 3. – С. 22–31.