
Ekosistemy, 31: 143–154 (2022) https://ekosystems.cfuv.ru
УДК 582.59:581.162.3 (477.75)
Некоторые антэкологические особенности орхидеи Neotinea tridentata в Крыму: пространственное размещение, параметры и цветовая гамма соцветий
Курамова В. В., Иванов С. П., Сволынский А. Д.
Крымский федеральный университет имени В. И. Вернадского Симферополь, Республика Крым, Россия
viktoriya.bekirova@ya.ru; spi2006@list.ru; svolinskiy@gmail.com
Приводятся сведения о характере распространения Neotinea tridentata на Крымском полуострове в целом, пространственном размещении в отдельных местообитаниях, строении и структуре соцветий, разнообразии окраски цветков, а также о некоторых других антэкологических особенностях, обеспечивающих необходимый уровень опыления и, соответственно, воспроизводство данного вида. Отмечается приуроченность вида к луговостепнвм сообществам и избегание типично степных. Показана неравномерность распределения Neotinea tridentata, как по территории Крыма, так и в отдельных местообитаниях. Мозаичное распространение вида в локальных местообитаниях приобретает характер крайней неравномерности – от плотных скоплений, в которых расстояние между цветущими особями измеряется 1–3 см, до разреженных, с расстоянием между особями в несколько десятков метров. Размещение особей в пределах отдельных местообитаний сильно отклоняется от нормального, представляя собой другой вид распределения – распределение «разломанного стержня» МакАртура. Численность и плотность особей в отдельных ценопопуляциях, как и параметры соцветий, сильно варьирует по годам. Представлены сведения о высоте и ширине соцветий, вариабельности и соотношении величин этих параметров, высоте цветоносов и количестве цветков в соцветиях. Представлены данные о последовательном изменении формы соцветия в период цветения и разнообразии соцветий в отношении плотности цветков. Показано, что увеличение плотности цветков на одной из сторон рыхлых соцветий может достигаться за счет нарушения радиальной симметрии в расположении цветков относительно вертикальной оси соцветия. Цветовая гамма цветков варьирует в широких пределах. Доминируют соцветия светло-фиолетово-розоватых оттенков. Реже встречаются темные фиолетово-розоватые формы и светлые – почти белые, кремовые или желтоватые. В некоторых случаях отмечено изменение насыщенности окраски цветков в процессе цветения соцветия.
Ключевые слова: орхидные, Neotinea tridentata, распространение вида, пространственное размещение особей, форма и структура соцветий, цветовая гамма цветков, Крымский полуостров.
ВВЕДЕНИЕ
Неотинея трехзубчатая – Neotinea tridentata (Scop.) R. M. Bateman, Pridgeon & M. W. Chase относится к одной из распространенных орхидей Крыма. В составе лугово-степной растительности она произрастает повсеместно в Горном и Предгорном Крыму, на части Южного берега Крыма (ЮБК), отмечена в Степной зоне (Каралларская и Осовинская степи на Керченском полуострове). Распространена локально в виде отдельных ценоопуляций, чаще состоящих из небольшого числа генеративных особей, но в некоторых местообитаниях образует скопления до 1000 генеративных особей (Красная книга…, 2015). Внесена в Красный список МСОП как вид, находящихся под угрозой исчезновения, в Красную книгу Российской Федерации (2008) со статусом «редкий вид», в Красную книгу Республики Крым (2015) с таким же статусом и в Красную книгу города Севастополя (2018) со статусом «сокращающиеся в численности».
Ареал вида охватывает юг и центр Европы, северную Африку, Малую Азию, Кавказ и Закавказье. Несмотря на широкое распространение и относительную многочисленность, о биологических особенностях этого вида известно немногое. В частности, данные по антэкологии вида ограничиваются сведениями о безнектарности цветков, опылителях и предположениями в отношении способа их привлечения (Müller, 1878; Voth, 1999a, 1992c; Salkowski, 2000; Berger, 2004, 2006a; Cozzolino et al., 2005; Claessens & Kleynen, 2011).
Некоторые особенности биологии и экологии Neotinea tridentata в Крыму приводятся в
ISSN 2414-4738 Published by V. I. Vernadsky Crimean Federal University, Simferopol
работах В. Назарова (1995) и С. Иванова с соавторами (Иванов и др. 2008, 2009а, 2009б), а также в монографиях (Kreutz et al., 2018; Фатерыга и др., 2019).
Цель работы – изучить особенности антэкологии орхидеи N. tridentata в Крыму, отражающие пространственное размещение вида в целом по Крыму и в пределах отдельных местообитаний, получить качественные и количественные характеристики пространственного распределения особей в пределах отдельных локалитетов, выявить особенности строения и структуры соцветий на разных стадиях цветения, оценить цветовую гамму цветков.
МАТЕРИАЛЫ И МЕТОДЫ
В качестве материала для изучения характера распространения N. tridentata в Крыму использованы данные, полученные в ходе экскурсий и экспедиционных выездов по Крыму, а также изучения литературных источников (Красная книга Республики Крым, 2015; Красная книга города Севастополя, 2018; Kreutz et al., 2018; Фатерыга и др., 2019).
Сбор материала по изучению характера пространственного распределения N. tridentata в локальных местообитаниях, а также по выявлению количественных и качественных характеристик соцветий и другим вопросам антэкологии проводили, начиная с 2000 года, в Предгорной зоне Крыма: в урочищах Бакла (рис. 1а), Аян (рис. 1б) и Мендер Крут-Ай (рис.
2а).
Выявление общего характера распределения генеративных особей в пространстве (тип дисперсии) проводили по Одуму (1975, стр. 265–267) в интерпретации Дайса (Dice, 1952). Количественную оценку степени сгруппированности особей осуществляли с использованием метрического классификатора, известного как «метод ближайшего соседа» (Харитонов, 2005), выявляя показатели, предложенные Кларком и Эвансом (Clark, Evans 1954), при этом сбор данных проводили по оригинальной методике. Наблюдатель приближался к группе цветущих особей и начинал измерять расстояние от первой попавшийся на его пути крайней особи до ближайшей в этой группе второй особи. Следующее измерение проводили от второй особи опять до ближайшей, разумеется, исключая предыдущую. Далее следовало измерение от третьей особи до ближайшей, исключая все предыдущие, и так далее. Измерения заканчивали, когда все особи группы были охарактеризованы на предмет оценки расстояния до ближайшей цветущей особи. Таким образом, сбор данных имитировал полет идеального опылителя, посещающего все соцветия в группе и выбирающего ближайшее соцветие каждый раз при очередном перелете от одного соцветия к другому. Нельзя не отметить, что, осуществляя такой «полет шмеля», наблюдатель имитировал идеальную ситуацию – неутомимость опылителя, его максимальную мотивированность на сбор провизии и достаточную наполненность цветков нектаром. В реальных условиях такая ситуация бывает крайне редко, но, на наш взгляд, такой метод сбора информации позволяет наиболее адекватно интерпретировать полученные данные, как в отношении стратегии растений по привлечению опылителей, так и стратегии опылителей в ходе сбора провизии.
Сбор дополнительных данных по выявлению пространственной структуры субпопуляций N. tridentata в отдельных местообитаниях проводили также на основе закладки трансект или картирования расположения цветущих особей.
Линейные параметры соцветий выявляли в полевых условиях, измеряя высоту и ширину соцветий линейкой. В условиях дефицита времени сбор материала проводили, фотографируя соцветия с приставленной к ним линейкой. Изменение формы соцветий по ходу их цветения, отклонения от симметрии в расположении цветков относительно оси соцветия и другие особенности антотаксиса фиксировали с помощью фотокамеры Canon EOS Rebel T2i с объективами Canon Zoom Lens EF-S 18-55 mm и Tamron SP Di AF 90 mm Macro 1:1. Цветовую гамму цветков в соцветиях оценивали по Таблице HTML-цветов
(https://vmirekraski.ru/prochee/tsveta-html).

Рис. 1. Местообитания Neotinea tridentata в Предгорьях Крыма
Остепенённые склоны массива Бакла (а) и склоны нижнего плато Чатыр-Дага в урочище Аян (b) – типичные варианты лугово-степной растительности в Крымских предгорьях.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Пространственная структура популяции и отдельных ценопопуляций Neotinea tridentata в Крыму. На территории Крыма N. tridentata отмечена в 120-ти отдельных местообитаниях (Фатерыга и др., 2019). Распространение вида на Южном берегу Крыма ограничено – вид не отмечен на значительном протяжении Крымского Южнобережья от Алушты до Судака, которое отличается относительной сухостью климата. Его

Рис. 2. Цветение Neotinea tridentata в урочище Мендер Крут-Ай
Две группы цветущих растений (а, b) и растение, цветущее одиночно (с); а – 14.05.21, b – 21.05.22, с – 23.05.21.
распространение в предгорьях Крыма, так же, видимо, определяется в основном режимом влагообеспечения – N. tridentata входит в состав исключительно лугово-степных фитоценозов, избегая типично степных склонов (рис.1, 2). Видимо, по этой же причине вид отсутствует в степной зоне Крыма. Исключение составляет единственное местообитание на Керченском полуострове, где вид встречается в небольшом числе локально на спускающихся к Азовскому морю северных прибрежных склонах Керченского мелкогорья к западу от села Осовины и в районе Генеральских пляжей (Караларская и Осовинская степи).
В отдельных местообитаниях N. tridentata также встречается в виде локальных группировок, число особей в которых может насчитывать от нескольких единиц до нескольких десятков, сотен и более цветущих особей. Наши многолетние наблюдения за изменением численности цветущих особей в отдельных локалитетах в Крыму показали, что количество и плотность особей в отдельных местообитаниях может существенно отличаться по годам, увеличиваясь в разы или снижаясь до единичных особей. В отдельные особенно засушливые сезоны, как например сезоны 2019 и 2020 годов в урочище Аян, нами было отмечены либо единичные, либо полное отсутствие цветущих особей N. tridentata.
Пространственное распределение цветущих особей N. tridentata наиболее полно исследовано нами в урочище Мендер Крут-Ай. Здесь на северо-западном склоне горы, давшей название урочищу, нами обнаружена ценопопуляция N. tridentata, включающая более сотни цветущих особей. Наибольшее скопление особей было обнаружено на участке целинного луга, протянувшегося полосой длиной около 100 и шириной 15–25 м, расположенного между полем (многолетняя залежь) и кустарниковыми зарослями. В юго-западном направлении этот участок лугово-степной растительности продолжался на расстояние еще 150 м, расширяясь местами до ширины 45 м. В этой части участка цветущие особи орхидеи встречались либо отдельными группами по 2–7 особей, либо одиночно (рис. 2).
В целом на территории, занятой данной ценопопуляцией N. tridentata, нами выделено три типа размещения цветущих особей: единичные особи на расстоянии 1–5 м друг от друга, единичные тесные группы по 2–7 особей на расстоянии 5–10 м друг от друга (и 1–7 см между особями в группе), а также большое скопление единичных особей в количестве более 400, расстояние между которыми варьировало от 5 до 230 см (рис. 3 и 4).
Характер распределения генеративных особей в пределах описанного скопления отражает рисунок 3.
Гистограмма распределения цветущих особей N. tridentata по расстоянию до ближайшего соседа на участке наибольшего скопления показывает, что более половины особей (51) находятся в среднем на расстоянии 32 см друг от друга. Одна четверть особей (19) – в среднем на расстоянии 86 см и только 1/10 особей – на расстоянии от 140 до 240 см. Среднее расстояние в скоплении – 57 см. Характер распределения особей по классам свидетельствует о том, что данное распределение не соответствует параметрам нормального. Уменьшение числа классов до 6 и сужение классового интервала до 40 см приводит это распределение к виду типичного распределения «разломанного стержня» МакАртура (MacArthur, 1957) (рис.
4а).
Неравномерность пространственного размещения цветущих особей N. tridentata ярко демонстрирует и рисунок 4b, из данных которого следует, что на отдельных 10-ти метровых отрезках вдоль трансекты, их число колеблется от 1 до 86.
Строение и структура соцветий, цветовая гамма цветков. Основные метрические показатели соцветий в двух ценопопуляциях Крыма (урочища Аян и Мендер Крут-Ай) в разные годы представлены в таблице 1. Из данных таблицы следует, что в одной и той же ценоопопуляции показатели разных лет могут существенно отличаться. В 2014 году по сравнению с 2010 в ценопопуляции в урочище Аян все показатели численно превосходили показатели 2010 года и были сравнимы с показателями субпопопуляции Мендер Крут-Ай 2022 года, за исключением высоты цветоноса.
Форма соцветия N. tridentata зависит от стадии цветения и количества цветков. На стадии зацветания первых цветков соцветие имеет четко выраженную пирамидальную форму (рис. 2а, 5а, 5b), независимо от общего количества цветков в соцветии (большая часть которых в обоих случаях пока находится в состоянии бутонов). В дальнейшем соцветие с небольшим числом цветков (не более 15–17) последовательно приобретает куполообразную форму (5f, 7d) и форму нечетко выраженного невысокого цилиндра, у которого высота может быть равна или даже быть меньше диаметра (рис. 5i, 7h).
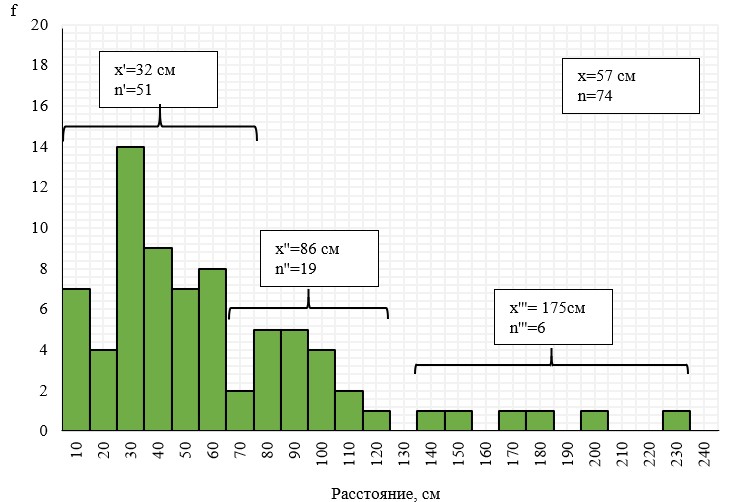
Рис 3. Гистограмма распределения цветущих особей Neotinea tridentata по расстоянию до ближайшего соседа на участке наибольшего их скопления (Мендер Крут-Ай, 2021 г.)
Классовый интервал 10 см. Средние величины расстояния подсчитаны отдельно для трех групп особей, находящихся друг от друга на расстоянии до 70 см, от 60 до 120 см и от 120 до 230 см.
b
а
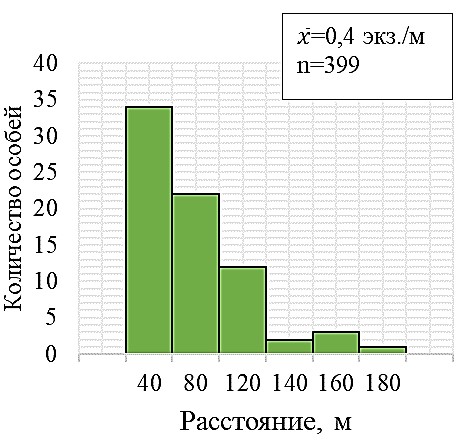
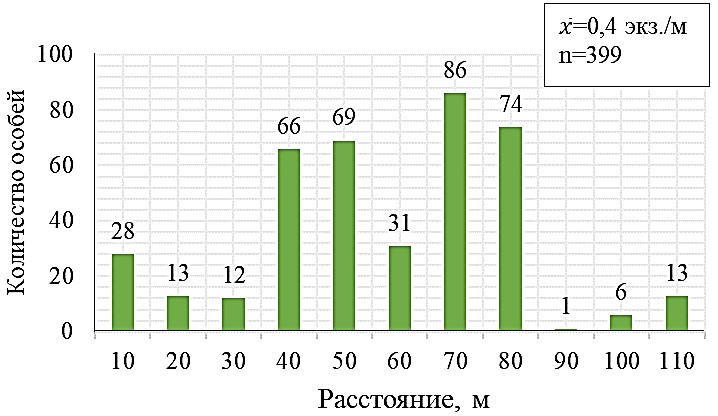
Рис. 4. Характер размещения цветущих особей Neotinea tridentata в скоплении
(Мендер Крут-Ай, 2021 г.)
а – гистограмма распределения цветущих особей по расстоянию до ближайшего соседа, классовый интервал 40 см; b – распределение особей вдоль линейной трансекты длиной 110 м.
Таблица 1
Параметры генеративных побегов Neotinea tridentata в двух субпопуляциях
Предгорного Крыма
| Пункт, год | N | Параметры цветочного побега и соцветия, x̄ ± σ | ||||
| Высота цветущего растения, см | Число цветков в соцветии | Высота соцветия, мм
(h) |
Диаметр соцветия, мм
(d) |
Соотношение
h/d |
||
| Аян, 2010 | 25 | 135,3 ± 20,9 | 19,0 ± 6,3 | 23,9 ± 4,7 | 23,8 ± 4,5 | 0,89 |
| Аян, 2014 | 25 | — | 28,0 ± 7,4 | 31,4 ± 7,5 | 26,8 ± 4,2 | 1,17 |
| Мендер Крут-Ай, 2021 | 30 | 155,7 ± 37,6 | 19 ± 6,4 | 38,2 ± 10,5 | 24,8 ± 5,1 | 1,54 |
| Мендер Крут-Ай, 2022 | 27 | 160,1 ± 30,3 | 27,3 ± 7,4 | 30,7 ± 9,2 | 26,7 ± 2,8 | 1,15 |
Примечание к таблице. N – число исследованных соцветий; x̄ ± σ – среднее значение и стандартное отклонение.
При большем числе цветков вначале также наблюдается пирамидальная (2а) и куполообразная форма, но последняя далее переходит в форму цилиндра с куполом (5с и 7а). Эта форма сохраняется, пока не расцветут все цветки, когда соцветие приобретает форму высокого цилиндра, высота которого определяется числом цветков (2b).
Нами отмечены различия соцветий по плотности размещения цветков в соцветии. В наиболее плотных соцветиях цветки максимально прижаты друг к другу, при этом, между цветками иногда нет даже небольшого свободного пространства (рис. 5 b, g; 7 a, d). Из-за этого некоторые цветки вынуждено меняют положение на наклонное (рис. 5 b и др.).
Иногда максимальные плотность цветков может достигаться и за счет нарушения радиальной симметрии цветков – смещением цветков в одну сторону (рис. 5 h, j, k, l).
Более детальный разбор особенностей антотаксиса N. tridentata будет представлен в следующей публикации.
Каждый цветок в соцветии цветет в течение 12–15 дней, после чего начинается процесс его увядания, а затем и высыхания (рис. 6 а, b, с).
Поскольку зацветание цветков в соцветии происходит не одновременно, а последовательно (начиная с нижних цветков), то и отцветание их происходит не одновременно. После нескольких дней нахождения соцветия в состоянии цветения всех цветков у нижних цветков начинают появляться первые признаки увядания (рис. 6a).
Процесс увядания цветка начинается раньше указанного выше срока, если произойдет его опыление. Уже через 1–2 дня после опыления начинается увядание цветка, и на 4–5 день цветок засыхает. В этом случае первые отцветшие цветки могут появиться в середине полностью цветущего соцветия (рис. 6b).
Плоды по мере созревания изменяют положение по отношению к оси соцветия, прижимаясь к оси и сближаясь друг с другом (рис. 6d и e в сравнении). Неопыленные цветки не меняют своего положения относительно оси соцветия (рис. 6f).

Рис. 5. Форма соцветий Neotinea tridentata
Стадии цветения: начало цветения (а); цветение половины цветков (b); цветение двух третей цветков (c); стадия полного цветения плотного (d) и рыхлого (g) соцветий с большим числом цветков; стадия полного цветения плотных (е и i) и рыхлого (f) соцветий с небольшим числом цветков. Нарушение радиальной симметрии в расположении цветков и достижение за этот счет большей их плотности:
шаровидное соцветие с одной (j) и другой (k) стороны; удлиненное соцветие с двух сторон (h и l).

Рис. 6. Плодоношение орхидеи Neotinea tridentata
Цветущие соцветия на стадии отцветания первых (а), большинства (b) и всех (c) цветков; вид соцветий в период созревания плодов (d–f).
Цветовая гамма окраски цветков N. tridentata в изученных локалитетах изменялась в широком диапазоне (рис. 7). Большинство соцветий имели цветки светло-фиолетовых оттенков. Более темные фиолетово-розоватые формы встречались редко. Также редки отклонения в светлую сторону, а очень светлые или почти белые формы встречались единично и не во всех местообитаниях. Наиболее редки кремовые или светло-желтые формы. В некоторых случаях отмечено изменение тона окраски цветков в процессе цветения соцветия (рис. 7а). На рисунке 7 представлены фото соцветий, сделанные в разное время суток. Фото а, b, c, d, е, h сделаны в интервале 11–13 часов дня, фото f, g – в интервале 15–16 часов. Во второй половине дня в связи с уменьшением высоты стояния солнца над горизонтом

Рис. 7. Цветовая гамма окраски цветков Neotinea tridentata
Цветки разной окраски в пределах одного соцветия (a): нижние цветки потеряли насыщенность окраски за время цветения (оттенок цвета цветков, сохранивших насыщенность – #cf74d6 по Таблице HTMLцветов). Цветовые формы соцветий: b – светло-розовато-фиолетовая (#c23bca); с – бело-фиолетовая (#f7e3ff); d – светло-фиолетовая (#e4a5f0); е – розовато-фиолетовая (#c380c4); f – светло-желтая
((#fcf6dd); g – кремовая (#f4dbd6); h – розовато-кремовая ((#e98bc0).
происходит сдвиг спектра солнечного света к более низким частотам (усиление желтоватых, а затем и красноватых оттенков). Фотографии, представленные на рисунке 7, не подвергались корректировке цветового баланса и, таким образом, представляют соцветия такими, какими их могут видеть пчелы в соответствующее время суток.
ЗАКЛЮЧЕНИЕ
Неотинея трехзубчатая (N. tridentata) относится к одной из наиболее распространенных и относительно многочисленных в Крыму видов орхидей. В то же время практически полное ее отсутствие в Степной зоне и большей части Южного берега Крыма свидетельствует о наличии некоторых неприемлемых условий для существования этого вида в некоторых районах. Но и на оставшейся, достаточно обширной территории обитания N. tridentata в Крыму, встречается крайне неравномерно. При этом очевидная приуроченность N. tridentata к лугово-степным вариантам растительности явно не достаточна для объяснения мозаичности распространения этой орхидеи в пределах, например, Предгорной зоны Крыма. Об особой требовательности к условиям обитания, узкой стенотопности вида говорит и своеобразная картина размещения особей в пределах отдельных местообитаний, которая соответствует особому типу распределения – распределению «разломанного стержня» МакАртура. Для выявления причин своеобразия пространственного размещения N. tridentata в Крыму необходимы дальнейшие исследования.
Численность и плотность особей N. tridentata в отдельных ценопопуляциях, как и параметры соцветий, сильно колеблются по годам, что, видимо, является реакцией растений на ухудшение или улучшение условий обитания в отдельные сезоны.
Размеры, форма и плотность цветков в соцветии также существенно варьируют. Полученные нами данные о последовательном изменении формы соцветия в период цветения, разнообразии соцветий в отношении количества и плотности цветков, а также широком спектре их окраски, требуют тщательного анализа в плане выявления значения этого разнообразия для привлечения опылителей и выявления механизма их аттрактации. Этим вопросам будет посвящена следующая наша публикация.
Список литературы
Иванов С. П. Фатерыга А. В., Тягнирядно В. В. Сравнительная оценка эффективности опыления орхидей в урочище Аян // Бюллетень Никитского ботанического сада. – 2008. – Вып. 97. – С. 10–14.
Иванов С. П., Холодов В. В., Фатерыга А. В. Орхидеи Крыма: состав опылителей, разнообразие систем и способов опыления и их эффективность // Ученые записки Таврического национального университета им. В. И. Вернадского. Серия «Биология, химия». – 2009а. – Т. 22 (61), № 1. – С. 24–34.
Иванов С. П., Фатерыга А. В., Фатерыга В. В. Эффективность опыления орхидей (Orchidaceae), цветущих одиночно и группами // Бюллетень Никитского ботанического сада. – 2009б. – Вып. 98. – С. 22–26.
Красная книга города Севастополя. – Главное управление природных ресурсов и экологии города Севастополя. – Калининград: Севастополь: ИД «РОСТ-ДОАФК», 2018. – 432 с.
Красная книга Республики Крым. Растения, водоросли и грибы / [Отв. ред. д.б.н., проф. А. В. Ена и к.б.н. А. В. Фатерыга]. – Симферополь: ИТ «АРИАЛ», 2015. – 480 с.
Красная книга Российской Федерации (растения и грибы) / Министерство природных ресурсов и экологии РФ; Федеральная служба по надзору в сфере природопользования; РАН; Российское ботаническое общество; МГУ им. М. В. Ломоносова [Отв. ред. Л. В. Бордунов, В. С. Новиков]. – М.: Товарищество научных изданий, 2008. – 855 с.
Назаров В. В. Репродуктивная биология орхидных Крыма: автореф. дис. … кад. биол. наук: 03.00.05 Ботаника. – Санкт-Петербург, Ботанический институт им. В. Л. Комарова, 1995. – 26 с.
Одум Ю. Основы экологии. – М.: Мир, 1975. – 740 с.
Фатерыга А. В. Ефимов П. Г., Свирин С. А. Орхидеи Крымского полуострова. – Симферополь: ИТ «АРИАЛ», 2019. – 224 с. Харитонов С. П. Метод «ближайшего соседа» для математической оценки распределения биологических объектов на плоскости и на линии // Вестник Нижегородского университета. Серия биология. – 2005. – № 1. – С. 213–221.
Berger L. Observations sur le comportement de quelgues pollinisateurs d’orchidees // L’Orchidophile. – 2004. – Vol. 35, N 1. – P. 19–35.
Berger L. Quelques notions de base sur la pollinisation orchidees // L’Orchidophile. – 2006. – Vol .37, N 3. – P. 183– 202.
Claessens J., Kleynen J. The flower of the European orchid. Form and function. – Voerendaal; Stein, 2011. – 439 p.
Clark P. J., Evans F. C. Distance to nearest neighbour as a measure of spatial relationships in populations // Ecology. – 1954. – Vol. 35, N 4. – P. 445–453.
Cozzolino S., Schiestl F. P., Muller A., Dе Castro O., Nardella A. M., Widmer A. Evidence for pollinator sharing in Mediterranean: absence of pre-mating barriers? // Proceedings of the Royal Society of London. Series B, Biological Sciences. – 2005. – Vol. 272. – P. 1271–1278.
Dice L. R. Measure of spaсing between individuals within a population // Contributions from the Laboratory of Vertebrate Biology, University of Michigan. – 1952. –Vol. 55. – P. 3-6.
Kreutz C. A. J., Fateryga A. V., Ivanov S. P. Orchids of the Crimea (description, pattern of life, distribution, threats, conservation and iconograhy). – Steenwijk: Kreutz Publishers, 2018. – 576 p.
MacArthur R. H. On the relative abundance of bird species // Proc. Nat. Acad. Sci. USA – 1957. – Vol. 45, N 3. – P. 293–295.
Müller H. Weitere Beobachtungen Uber Befruchtung der Blumen durch Insekten // Verhandlungen des Naturhistorischen Vereines der Preussischen Rheinlande und Westphalens. – 1878. – Vol. 35. – P. 272–291.
Salkowski H.-E. Die Erdhummel (Bombus terrestris) als Besucher und Bestauber von Orchideenbluten. Biobachtungen an drei Beispielen // Berichte aus den Arbeitskreisen Heimische Orchideen. – 2000. – Vol. 17, N. 2. – P. 140–141.
Voth W. Bestaubungs biologische Beobachtungen an Orchis tridentata Scop. – Orchidea. – 1992. – 43 (6). – P. 280– 283.
Voth W. Lebensgeschichte und Bestauber der Orchideen am Beispiel von Niederosterreich // Stapfia. – 1999. – Vol.
65. – P. 1–257.
Kuramova V. V., Ivanov S. P., Svolynsky A. D. Some antecological features of the orchid Neotinea tridentata in the Crimea: spatial distribution, parameters and colors of inflorescences // Ekosistemy. 2022. Iss. 31. P. 143–154.
The article gives information on the distribution pattern of Neotinea tridentata in the Crimean Peninsula and its spatial distribution in some habitats, the structure of the inflorescence, the variety of flower colors, as well as some other antecological features that ensure the necessary level of pollination, and, consequently, the reproduction of the species. The species area coincides with meadow communities and avoids steppe ones. The uneven distribution of Neotinea tridentata is revealed both in the territory of Crimea and in certain habitats. The mosaic distribution of the species in local habitats is extremely irregular, ranging from dense aggregations in which the distance between flowering individuals is 1–3 cm, to sparse ones with a distance of several tens of meters between individuals. The distribution of individuals within some habitats deviates strongly from the normal one, presenting MacArthur’s “broken stick” distribution. The number and density of individuals in separate coenopopulations and the inflorescence parameters vary significantly from year to year. The information is presented on inflorescence length and width, the variability and ratio of these parameters, pedicel height, and the number of flowers in the inflorescence. The article provides data on the consecutive change in the shape of inflorescence during the flowering period and, moreover, on the variety of inflorescences regarding the density of flowers. It is shown that an increase in flower density on one side of an inflorescence can be achieved by breaking the radial symmetry in the arrangement of the flowers about the vertical axis of the inflorescence. The color range of the flowers varies widely. Inflorescences of light violet-pinkish shades dominate. Darker violet-pinkish forms and lighter forms (almost white, cream or yellowish) are less common. In some cases, a change in the color saturation of flowers during flowering process is registered.
Key words: orchids, Neotinea tridentata, spatial distribution, form and structure of inflorescences, color range of flowers, Crimean Peninsula.
Поступила в редакцию 10.10.22
Принята к печати 21.11.22
