
Экосистемы, 33: 38–63 (2023) https://ekosystems.cfuv.ru
![]()
УДК: 595.371-15(262.5+262.54)
Институт биологии южных морей имени А. О. Ковалевского РАН Севастополь, Россия
VGrintsov@gmail.com
На основе анализа морфологии, биологии, этологии и экологии 141 вида амфипод Черного и Азовского морей выделено пять экологических групп: трубкожилы, перфораторы, комменсалы, кривофилы (от греческого «kryvo»
– прятаться, «phileо» – любить) и мезопсаммофилы (от греческого «meso» – внутри, «psammo» – песок, «phileo» – любить). В пределах данных пяти экологических групп обозначены экоморфы и жизненные формы. У кривофилов выделена капреллидная экоморфа (семейство Caprellidae) и 9 жизненных форм, названных по таксонам, входящим
- них: хиалидная (род Hyale), дексаминидная (род Dexamine), гаммаридная (род Gammarus), эдицеротидная (семейство Oedicerotidae), мегалуропиидная (род Megaluropus), талитридная (семейство Talitridae), харпиниидная (род Harpinia), стенотоидная (род Stenothoe), лизианассидная (семейство Lisianassidae). Жизненные формы всех 5 экологических групп не ограничены таксонами, по которым они названы. Трубкожилы разделены на ампелисцидную (род Ampelisca), ампитоидную (род Ampithoe) и монокорофиидную (род Monocorophium) жизненный формы. У перфораторов выделена одна жизненная форма – бианколинидная (род Biancolina). У комменсалов обозначены три жизненных формы: кардиофилидная (род Cardiophilus), коломастигидная (род Colomastix) и леукотоидная (род Leucothoe). У мезопсаммофилов выделены две жизненных формы – батипореидная (род Bathyporeia) и маеотикоидная – вид Pontogammarus maeoticus (Sovinskij, 1894). В настоящей статье анализируется экологические группы, экоморфы и жизненные формы, а также их распределение по сообществам, биотопам, средам обитания или экотопам в соответствие с морфологическими идиоадаптациями амфипод, а также этологией, экологией и биологией этих ракообразных.
Ключевые слова: Amphipoda, экоморфы, экологические группы, жизненные формы, Черное море, Азовское
море.
ВВЕДЕНИЕ
Экологические группы (Димо, Келлер, 1907), экоморфы (Алеев, 1980) и жизненные формы (термин введен А. Гумбольтом в 1806 году, в зоологии одним из первых в современном значении приведен А. Н. Формозовым (1929)) живых организмов формируются
- процессе эволюции под влиянием той или иной среды обитания, в том числе через идиоадаптации, определяющие успех выживания организмов в предпочитаемых экотопах, биотопах, сообществах. Идиоадаптации не приводят к общему повышению уровня организации организмов как ароморфозы, но преобразуют частные изменения функции органов (Северцов, 1949).
- данной работе понятие «экологическая группа» описывается следующей формулировкой: «экологи́ческая гру́ппа — совокупность видов, характеризующаяся сходными потребностями в величине какого-либо экологического фактора и возникшими в результате его воздействия в процессе эволюции сходными анатомо-морфологическими и иными признаками, закрепившимися в генотипе». Для описания экоморф взята формулировка: «экоморфа – целостная система взаимообусловленных эколого-морфологических адаптаций, определяющая общую конструкцию тела организма в соответствие с конкретным направлением эволюции вида в условиях конкретного биотопа» (Алеев, 1980). Для описания жизненных форм использовали формулировку: «жизненная форма – внешний облик растений или животных, как систематически близких, так и далеких, отражающий их приспособленность к условиям среды» (Дедю, 1990).
ISSN 2414-4738 Published by V. I. Vernadsky Crimean Federal University, Simferopol
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
Цель данных исследований – проанализировать морфологические идиоадаптации, определяющие экологические группы, экоморфы и жизненные формы амфипод Черного и Азовского морей, а также распределение этих категорий по сообществам, биотопам, средам обитания или экотопам в соответствие с их морфологическими идиоадаптациями, особенностями поведения, биологии и экологии.
МАТЕРИАЛ И МЕТОДЫ
На основе результатов многолетних исследований морфологии, экологии, этологии и биологии 141 вида амфипод Черного и Азовского морей (Булычева, 1957; Мордухай-Болтовской и др., 1969; Василенко, 1974; Грезе, 1977, 1985; Маккавеева, 1979; Киселева, 1981; Petrescu, 1998; Sezgin et al, 2001; Гринцов, 2009, 2010, 2011а, 2011б, 2016, 2017, 2021, 2022а, 2022б, 2022 в; Grintsov, 2009, 2010, 2014, 2018, 2021а, 2021б; Grintsov, Sezgin, 2011; Uzunova, 2012; Kudrenko, 2016) выделялись экологические группы, экоморфы и жизненные формы амфипод этих водоемов.
Для обозначения данных категорий были использованы морфологические идиоадаптации, выработанные амфиподами в ответ на наиболее критические условия той среды (или сред) обитания, которую (или которые) эти амфиподы предпочитают.
Фотографии габитусов и деталей тела взрослых самцов и самок сделаны с использованием микроскопа Hitachi SU 3500. Контурные зоологические рисунки ряда видов взяты из литературных источников.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Рассмотрим особенности выделенных категорий. Весь последующий анализ идиоадаптации и на их основе обозначенных категорий, рассматривается по сравнению с гаммаридной жизненной формой (сем. Gammaridae: рода Gammarus, Echinogammarus, Chaetogammarus и др.), предположительно более древней, поскольку известны окаменелые останки амфипод, близких по морфологии к этому семейству (Arfianti et аl., 2018). Кроме того, представители данной жизненной формы менее специализированы, что условно указывает на это семейство как на предковую группу (Гурьянова, 1951). В качестве анализируемых морфологических элементов рассматриваются не любые морфологические элементы, а идиоадаптации, способствующие выживанию на фоне тех экстремальных условий среды обитания (или сред обитания), к которым эти экоморфы и жизненные формы наиболее приспособлены.
Рассмотрим выделенные категории. Анализировали только идиоадаптации в связи с определенными условиями среды обитания. Название элементов морфологии здесь и далее даны в соответствие с принятыми для этой группы (Гурьянова, 1951; Barnard, Karaman, 1991). Группа трубкожилов характеризуется способностью строить трубки из «паутинного шелка» (Гурьянова, 1951). От других амфипод их отличает ряд следующих признаков: коксальные пластинки слабо развиты (функция защиты брюшной части тела ослаблена, боковые щиты не выражены), оостегиты самок (выводковые пластинки) узкие и длинные, не образующие сплошной камеры (Гурьянова, 1951). К этим признакам можно добавить несколько особенностей. Тело часто более округлое в поперечном сечении или сплющенное в спино-брюшном направлении. Кутикула в спинной части сегментов тела без зубцов или шипов (отсутствует дополнительная функция защиты тела от хищников); переоподы вооружены «паутинными» железами и поэтому определенные членики утолщены.
- трубкожилов, зарегистрированных в Черном и Азовском морях, можно обозначить три жизненных формы (рис. 1).
Тело амфипод монокорофиидной жизненной формы (рис. 1 А) уплощено сверху вниз; четвертые членики стебельков 2-й пары антенн у самцов вооружены зубцами (имеет место
39
Гринцов В. А.
![]()

Рис. 1. Внешний вид представителей монокорофиидной (А) ампитоидной (Б) и ампелисцидной жизненных форм трубкожилов
- – Monocorophium insidiosum (Crawford, 1937); Б – Ampithoe ramondi Leach, 1814; В – Ampelisca diadema (Costa, 1853). Масштабная литейка 1 мм.
добавление функции по (Северцов, 1949), а именно функции захвата, рис. 1 А, «А 2, зубцы»). Гнатоподы обеих пар с рядами длинных перистых щетинок, расположенных в виде ловчей сети (рис. 2 А, пища собирается по типу фильтрации).
Коксальные пластинки не смыкаются, слабо развиты (рис. 1 А, функция защиты брюшка
- боков ослаблена). Плеоподы короткие (рис. 2 Б, указаны стрелкой) не вытянуты как у других групп амфипод, что возможно отражается на эффективности плавания. Уроподы 3-й пары короткие и широкие, с почти округлыми ветвями (рис. 2 В, указаны стрелкой), без крючков или развитых шипов, что указывает на отсутствие использования их как зацепа за какие-либо предметы. Особи монокорофиидной формы избегают прибойных участков, предпочитая места с накоплением детрита – бухты, лагуны, эстуарии рек. В Черном и Азовском морях в эту форму входят следующие рода: Chelicorophium (7 видов), Corophium (2), Monocorophium
(2), Medicorophium (1), Crassicorophium (1).
Тело амфипод ампитоидной жизненной формы (рис. 1 Б) сжато с боков или близко к округлому в поперечном сечении. Коксальные пластинки 1–4 хорошо развиты и образуют щит, закрывающий брюшную часть с боков. Антенны 1-й и 2-й пар выполняют у одних видов сенсорную функцию, у других к ней добавляется функция собирания частиц с помощью плотных групп щетинок. Когти переопод морфологически однотипны, в виде крючков, часто дополнительно дистальный конец переона усилен шипами или зубцами рядом с креплением когтя (рис. 2 Г, указаны стрелками, функция зацепа за субстрат усилена). Иногда и когти усилены дополнительными шипами, например, у Ericthonius difformis (Spence Bate, 1857). Базиподиты 6-й и 7-й пар по форме однотипны. Уроподы 3-й пары у одних видов по типу гаммаридной формы, но ветви менее вооружены и менее развиты (рис. 2 Д, указаны стрелками). У других морфологически другие: стебельки короткие, ветви с крючками или мощными шипами (рис. 2 Ж, крючок указан стрелкой), что функционально выражается в смене роли рулей при плавании (роль рулей при плавании по (Barnard, Karaman, 1991), характерных для гаммаридной формы (рис. 10 К), на роль зацепа за элементы окружающей среды. Следует уточнить, что ветви 3-й пары уропод по типу рулей вытянутые, уплощенные или округлые в сечении, снабженные щетинками и шипами (рис. 10 К). Ветви уропод 3-й пары по типу зацепа короткие, конические, вооруженные развитыми зубцами, шипами и/или щетинками преимущественно терминально (рис. 2 Ж). Амфиподы ампитоидной жизненной формы предпочитают скопления водорослей в перифитоне и на дне. В Черном и Азовском морях в эту форму входят рода Ampithoe (1 вид), Cheirocratus (1), Cymadusa (1), Ericthonius
(3), Jassa (3), Pleonexes (1), Plumulojassa (1), Megamphopus (1), Photis (1), Microdeutopus (5), Microprotopus (2), Centraloecetes (1), Leptocheirus (1).
-
- амфипод ампелисцидной жизненной формы (рис. 1 В) глаза рудиментарны, в виде отдельных маленьких линз (функция зрения ослаблена). Коксальные пластинки 1–4 хорошо развиты (функция защиты брюшка усилена с боков, выражены боковые щиты). Переоподы 4-й пары с длинным тонким когтем (рис. 2 З, коготь указан стрелкой) и рядами развитых
40
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
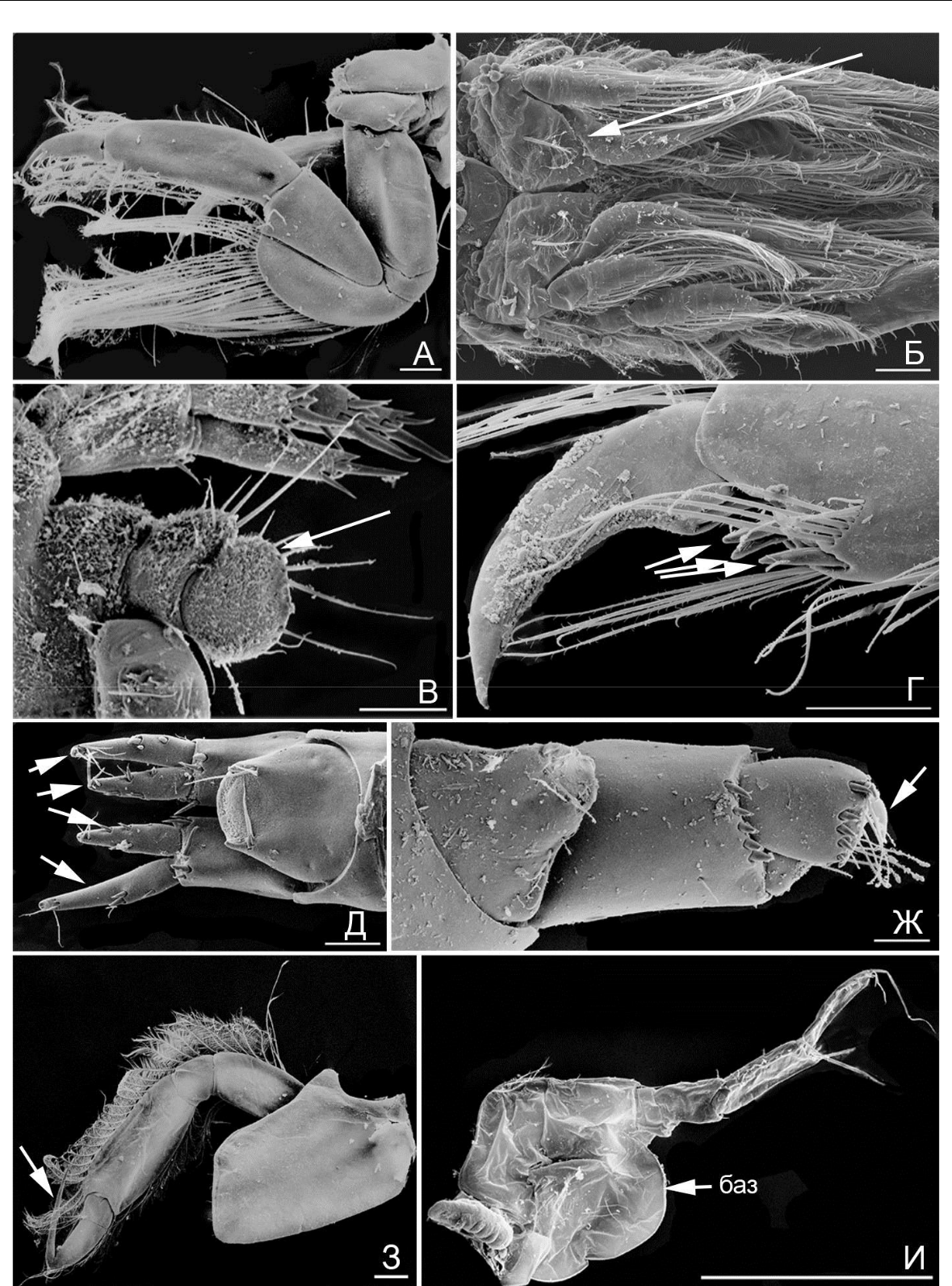
Рис. 2. Элементы морфологии амфипод
- – гнатопод 2, M. insidiosum; Б – плеоподы (стрелка), M. insidiosum; В – уросом и уропод 3 (стрелка), M. insidiosum; Г – A. ramondi, переопод 7, коготь и дистальная часть проподуса (стрелкой указаны шипы); Д – M. gryllotalpa, телсон и уроподы 3 (ветви указаны стрелкой); Ж – A. ramondi, тельсон и уропод 3; (стрелкой указан зубец); З – A. sevastopoliensis, переопод 4 (стрелкой указан коготь); И – A. diadema, переопод 6 («баз» – базиподит). Масштабная линейка 0,1 мм.
41
Гринцов В. А.
![]()
щетинок различного типа (простые, перистые, гребнистые), что указывает на использование этой пары конечностей для чистки тела и собирания частиц, возможно для собирания пищи. Базиподиты 6-й и 7-й пар переопод разной формы (рис. 2 И «баз»; рис. 3 А, «баз»). Когти переопод 6-й пары с группой дополнительных шипов (рис. 3 Б, указаны стрелками, функция захвата усилена, возможно добавление функции распределения «шелка» при постройке трубок).
Когти переопод 7-й пары каплевидной формы (рис. 3 А «ког», смена функции захвата на функцию упора). Уроподы 3-й пары с вытянутыми ветвями (рис. 3 В, ветви указаны стрелками), по типу рулей, но шипы и щетинки более редкие, чем у представителей гаммаридной жизненной формы. Особи ампелисцидной формы обитают почти исключительно среди детрита на рыхлом грунте. В Черном и Азовском морях в эту форму входит род Ampelisca (4 вида).
-
- амфипод-перфораторов выделена одна бианколинидная жизненная форма, внешний вид которой представлен на рис. 4.
Тело амфипод бианколинидной жизненной формы в поперечном сечении близко к округлому (рис. 4 А, Б); голова полушарообразная (рис. 4 А). Коксальные пластинки слабо развиты (рис. 4 А, Б, функция защиты брюшка ослаблена, за счет уменьшения коксальных пластинок, боковые щиты не выражены, функцию защиты при этом берут на себя ходы внутри перфорируемого объекта). Когти и проподальные членики вблизи когтей у некоторых видов (напр. B. algicola) могут быть усилены дополнительным вооружением (рис. 3 Г, шипы указаны стрелкой, функция зацепа усилена за счет усиления вооружения проподуса вблизи крепления когтя). Уроподы 3-й пары дополнительно вооружены (рис. 3 Д, шипы указаны стрелкой, функция зацепа усилена за счет добавления шипов или зубцов). Особи обитают либо внутри столонов крупных бурых макрофитов (B. algicola), либо в ходах внутри древесины (C. terebrans). В Черном и Азовском морях в эту форму входят два вида –
B. algicola, C. terebrans.
Комменсалы были разделены на 3 жизненные формы (рис. 5).
Тело амфипод коломастигидной жизненной формы в поперечном сечении почти округлое (рис. 5 А). Антенны обоих пар короткие, жгутики слабо развиты (рис. 5 А, сенсорная функция ослаблена за счет снижения числа члеников жгутиков, вооруженных чувствительными органами). Коксальные пластинки узкие, не закрывают полностью брюшную часть переона с боков (рис. 5 А, функция защиты брюшка с боков ослаблена, не выражены боковые щиты). Переоподы близкие по длине, их когти короткие (функция зацепа когтями за предметы ослаблена). Ветви 3-й пары уропод вытянуты, сужаются дистально, по типу рулей, но с более редкими шипами и щетинками, чем у представителей гаммаридной формы (напр. у Gammarus, Echinogammarus и пр.). В Черном море в эту жизненную форму входит один вид – C. pusilla.
Тело амфипод леукотоидной жизненной формы (рис. 5 Б) сжато с боков. Антенны 1-й и 2-й пар почти равны по длине, со слабо развитыми жгутиками (рис. 3 Б, сенсорная функция ослаблена за счет снижения числа члеников жгутиков, вооруженных чувствительными органами). Коксальные пластинки 1–4 хорошо развиты, образуют боковые щиты. Гнатоподы обоих пар (особенно 1-й) с характерным развитым выростом на карпальном членике (рис. 5 Б, функция захвата предметов элементами гнатопод усилена). Когти переопод небольшие и слабо загнутые (рис. 5 Б, функция захвата субстрата ослаблена). Уроподы с ветвями по типу рулей, сужающимися дистально, вооружение шипами и щетинками относительно гаммаридной формы значительно редуцировано (вероятно, ослаблена функция рулей при плавании). В Черном море (исключительно прибрежье Турции) в эту форму входит L. spinicarpa.
Тело амфипод кардиофилидной жизненной формы (рис. 5 В) сжато с боков. Антенны 1-
- пары длиннее антенн 2-й пары (рис. 5 В, сенсорная функция антенн 1-й пары усилена за счет увеличения числа члеников жгутиков с чувствительными органами). Гнатоподы 1-й и 2-й пар
42
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей

Рис. 3. Элементы морфологии амфипод
- – A. sevastopoliensis, переопод 7 («баз» – базиподит; «ког» – коготь); Б – A. sevastopoliensis, переопод 6, коготь (стрелкой указаны шипы); В – A. sevastopoliensis, уропод 3 (стрелкой указаны ветви); Г – B. algicola, переопод 5, коготь и дистальная часть проподуса (стрелкой указан шип); Д – B. algicola, уропод 3 и тельсон (стрелкой указаны шипы на ветвях уроподов 3); Ж – C. baeri, гнатопод 1; З – C. baeri, гнатопод 2; C. baeri, уросом, уроподы (стрелками указаны ветви уропод 3). Масштабная линейка: А, В,
Д-И – 0,1 мм; Б, Г – 0,01 мм.
43
Гринцов В. А.
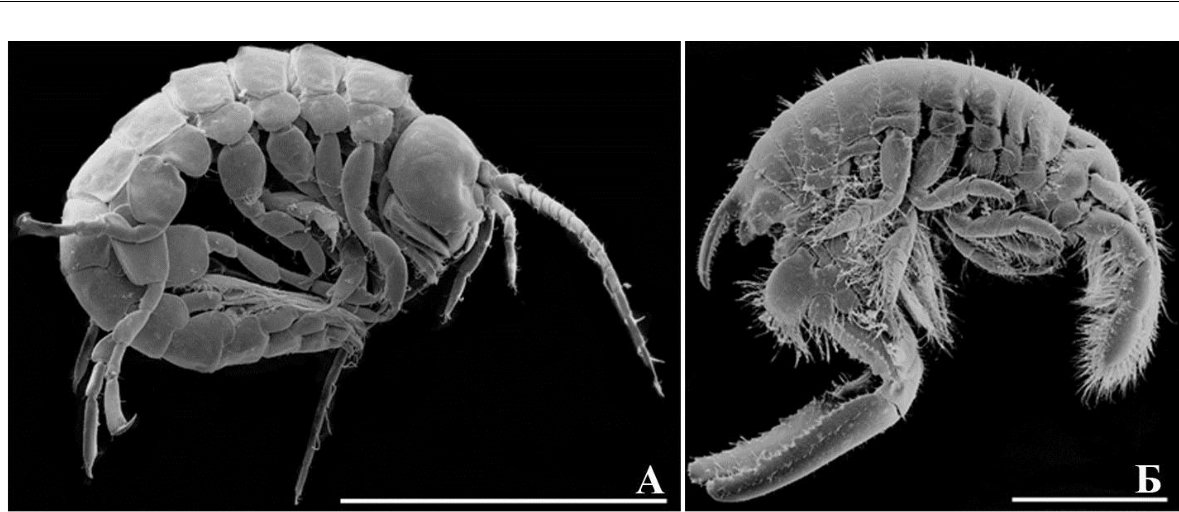
Рис. 4. Внешний вид особей бианколинидной жизненной формы
- – самка Biancolina algicola Della Valle, 1893; Б – самец Chelura terebrans Philippi, 1839. Масштабная линейка 1 мм.
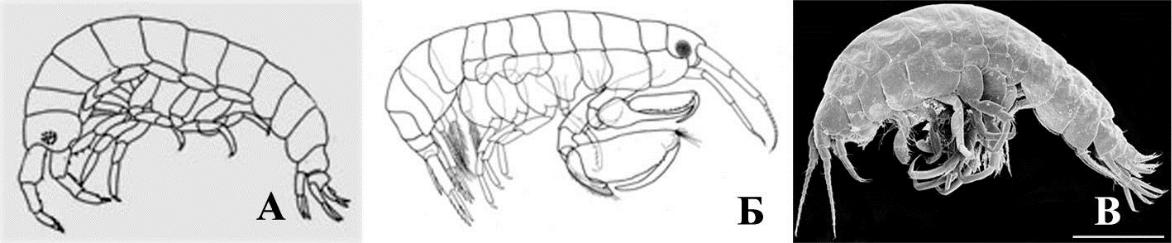
Рис. 5. Внешний вид представителей жизненных форм комменсалов: коломастигидной (А), леукотоидной (Б), кардиофилидной (В)
- – Colomastix pusilla Grube, 1861 (рис. по Barnard & Karaman, 1991); Б – Leucothoe spinicarpa (Abildgaard, 1789), (рис. по Krapp-Schickel, 1989); Г – Cardiophilus baeri G. O. Sars, 1896. Масштабная линейка 1 мм.
слабо развиты, разной формы (рис. 3 Ж, З, функция захвата предметов ослаблена). Коксальные пластинки 1–4 хорошо развиты (защита брюшной части переона с боков усилена). Уроподы 3-й пары с вытянутыми ветвями, но значительно меньше относительно тела и слабее вооружены, чем у гаммаридной жизненной формы (рис. 3 И, ветви указаны стрелками). Форма отмечена как комменсал двустворчатых моллюсков семейства Cardiidae, но также регистрируется в сообществах бентоса вне этих моллюсков. В Черном и Азовском морях эта форма представлена двумя видами рода Cardiophilus.
Мезопсаммофилы – амфиподы, зарывающиеся в песок (от греческого «meso» – внутри, «psammo» – песок, «phileo» – любить) были разделены на две жизненные формы (рис. 6).
Тело амфипод батипореидной жизненной формы (рис. 6 А) сужается от абдомена к голове (функция проникновения в песок облегчена). Между 1-м и 2-м члениками стебелька 1-
- пары антенн имеется перегиб на 90°, а сам дистальный участок 1-го членика с усиленным выростом (рис. 7 А, защита жгутика антенны усилена, указано стрелкой).
Коксальные пластинки 1–5 хорошо развиты, перекрываются, с венчиками длинных щетинок по краям (рис. 6 А, функция защиты брюшка с боков усилена, выражены боковые щиты). Гнатоподы 2-й пары без когтя, обильно снабжены группами длинных щетинок, что указывает на их собирающую функцию, но не на захват пищи обычным способом с помощью когтя (рис. 7 Б). Членики переопод значительно расширены, роющего типа (рис. 7 В, функция зарывания в песок усилена), вооружены множеством шипов и щетинок; когти переопод массивные, не загнутые (рис. 7 Г, указаны стрелкой, смена функции захвата на таковую
44
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
зарывания). Уроподы 3-й пары по типу рулей, с одной, но сильно развитой ветвью (рис. 7 Д, указана стрелкой), другая ветвь редуцирована до небольшой пластинки, менее вооружена щетинками и шипами по бокам, чем у представителей гаммаридной формы. Обитает ниже прибойной зоны, преимущественно глубже 10 м (Грезе, 1985). В Черном и Азовском морях отмечен один вид B. guilliamsoniana.
Амфиподы маеотикоидной жизненной формы (рис. 6 Б) с массивными антеннами, обильно вооруженными плотными группами щетинок (рис. 7 Ж), членики жгутиков антенн четковидные, массивные (функция проникновения в песок усилена, выражена функция защиты ротовых органов от песчинок). Коксальные пластинки 1–4 хорошо развиты, перекрываются, снабжены по свободным краям (вентрально, антеро-вентрально и постеро-вентрально) венчиками длинных щетинок (рис. 6, защитная функция брюшка с боков усилена, боковые щиты выражены). Гнатоподы и переоподы массивные, членики снабжены множеством длинных щетинок (рис. 7 З, функция зарывания в песок усилена), когти переопод развитые (рис. 7 З, функция захвата субстрата выражена). Уроподы 3-й пары по типу рулей, с развитыми ланцетовидными лопастями, обе пары лопастей (рис. 7 И, особенно наружная, указанная стрелкой) снабжены густой щеткой длинных перистых щетинок. В Черном и Азовском морях в маеотикоидную форму входит два рода и два вида понто-каспийских амфипод – Pontogammarus maeoticus (Sowinskyi, 1894) и Euxinia weidemanni (G. O. Sars, 1896).

Рис. 6. Внешний вид представителей батипореидной (А) и маеотикоидной (Б) жизненных форм мезопсаммофилов
- – Bathyporeia guilliamsoniana (Bate, 1857); Б – Pontogammarus maeoticus (Sowinskyi, 1894).
Масштабная линейка 1 мм.
Наконец, группа кривофилов (от греческого «kryvo» – прятаться, «phileо» – любить), предпочитает прятаться среди различных предметов (не используя указанные выше для других групп способы) – под камнями, среди водорослей, двустворчатых моллюсков, гидроидных полипов и пр. В данную группу входит большинство таксонов амфипод Черного
- Азовского морей. К этой группе относится капреллидная экоморфа, отличающаяся от всех остальных амфипод Черного и Азовского морей (семейство Caprellidae) и 9 жизненных форм. Амфиподы всех других экологических групп и оставшиеся амфиподы гаммаридной экологической группы по отношению к капреллидной экоморфе условно можно отнести к гаммаридной экоморфе. Гаммаридная экоморфа названа по семейству Gammaridae, входящему в нее, поскольку известны окаменелые остатки близкие к этому семейству (Arfianti et. al, 2018). Кроме того, Gammaridae менее специализированы по выбору субстрата относительно многих других семейств (Гурьянова, 1951), что условно указывает на это семейство как на предковую группу. В дальнейшем гаммаридная экоморфа анализироваться не будет, поскольку в нее входят все остальные амфиподы Черного и Азовского морей (кроме Caprellidae), которые будут рассматриваться подробнее по жизненным формам.
Капреллидная экоморфа (рис. 8 А, Б) отличается от гаммаридной экоморфы палочковидным телом, олигомеризацией сегментов тела, цефализацией, редукцией придатков – переопод, плеопод, уропод, а также усилением проподального членика и когтя.
45
Гринцов В. А.
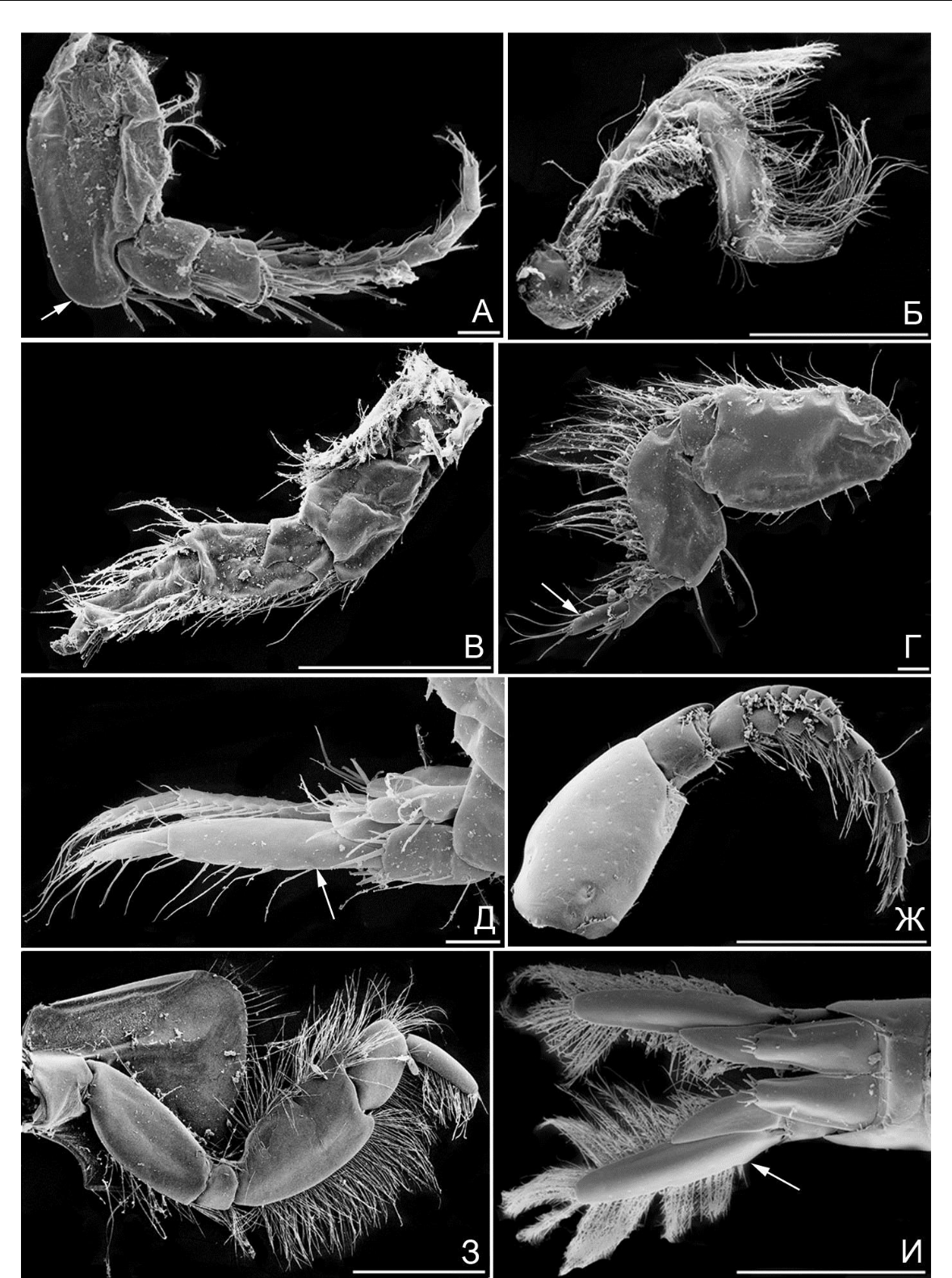
Рис. 7. Элементы морфологии амфипод
- – B. guiliamsoniana, антенна 1 (стрелкой указан выступ 1-го членика стебелька); Б – B. guiliamsoniana, гнатопод 2; В – B. guiliamsoniana, переопод 7; Г – B. guiliamsoniana, переопод 6 (стрелкой указан коготь); Д – B. guiliamsoniana, уроподы 3 (стрелкой указана наружная ветвь уропод 3); Ж – P. maeotica, антенна 1; З – P. maeotica, переопод 3; И – P. maeotica, уроподы 3 (стрелкой указана наружная ветвь). Масштабная линейка 1 мм.
46
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
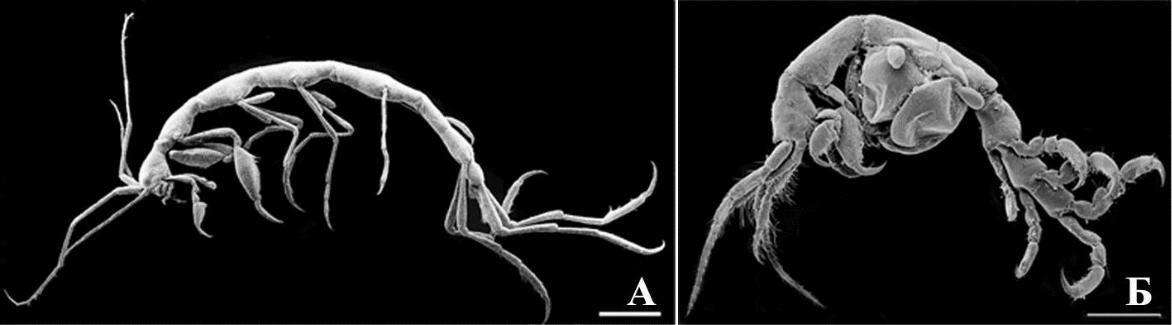
Рис. 8. Внешний вид представителей капреллидной экоморфы кривофилов
- – Phtisica marina Slabber, 1769; Б – Caprella liparotensis Haller, 1879. Масштабная линейка 1 мм.
Рассмотрим 9 жизненных форм кривофилов. Тело амфипод дексаминидной жизненной формы (рис. 9 А) с зубцами, либо валиками на спине (рис. 9 А, функция защиты от хищников усилена).

Рис. 9. Внешний вид представителей дексаминидной (А) хиалидной (Б) и гаммаридной жизненных форм кривофилов
- – Dexamine spinosa (Montagu, 1813); Б – Apohyale crassipes (Heller, 1866); В – Gammarus subtypicus Stock, 1866. Масштабная линейка 1 мм.
Капреллидная экоморфа обитает преимущественно на водорослях и гидроидных полипах. Несмотря на специализацию по среде обитания, эта экоморфа широко распространена из-за наличия водорослей почти во всех биотопах и сообществах перифитона
- бентоса. В Черном море к данной экоморфе относятся рода Caprella (6 видов), Phtisica (1), Pseudoprotella (1).
Антенны длинные или снабжены множеством чувствительных органов (рис. 10 А, Gammarellus), что указывает на усиление сенсорной функции. Гнатоподы однотипные по морфологии и сходные по размерам (рис. 10 Б, В). Коксальные пластинки 1–4 хорошо развиты, перекрываются (рис. 9 А, защита брюшка с боков усилена, боковые щиты выражены). Когти переопод развитые, загнутые (рис. 10 Г, указано стрелкой, функция захвата субстрата усилена). Уроподы 3-й пары с развитыми ланцетовидными ветвями (рис. 10 Д, ветви по типу рулей, указаны стрелками), однако вооружение менее обильно, по сравнению с представителями гаммаридной формы. Амфиподы дексаминидной формы характерны для водорослевых сообществ, преимущественно перифитона или различных скоплений водорослей, как в перифитоне, так и в бентосе. К дексаминидной жизненной форме в Черном
- Азовском морях относятся рода Apherusa (2 вида), Dexamine (3), Gammarellus (2), Tritaeta
(1), Nototropis (2).
Амфиподы эдицеротидной жизненной формы (рис. 11 А), с глазами, сдвинутыми к теменной части головы, или глаза отсутствуют (изменение или потеря функции зрения).
47
Гринцов В. А.
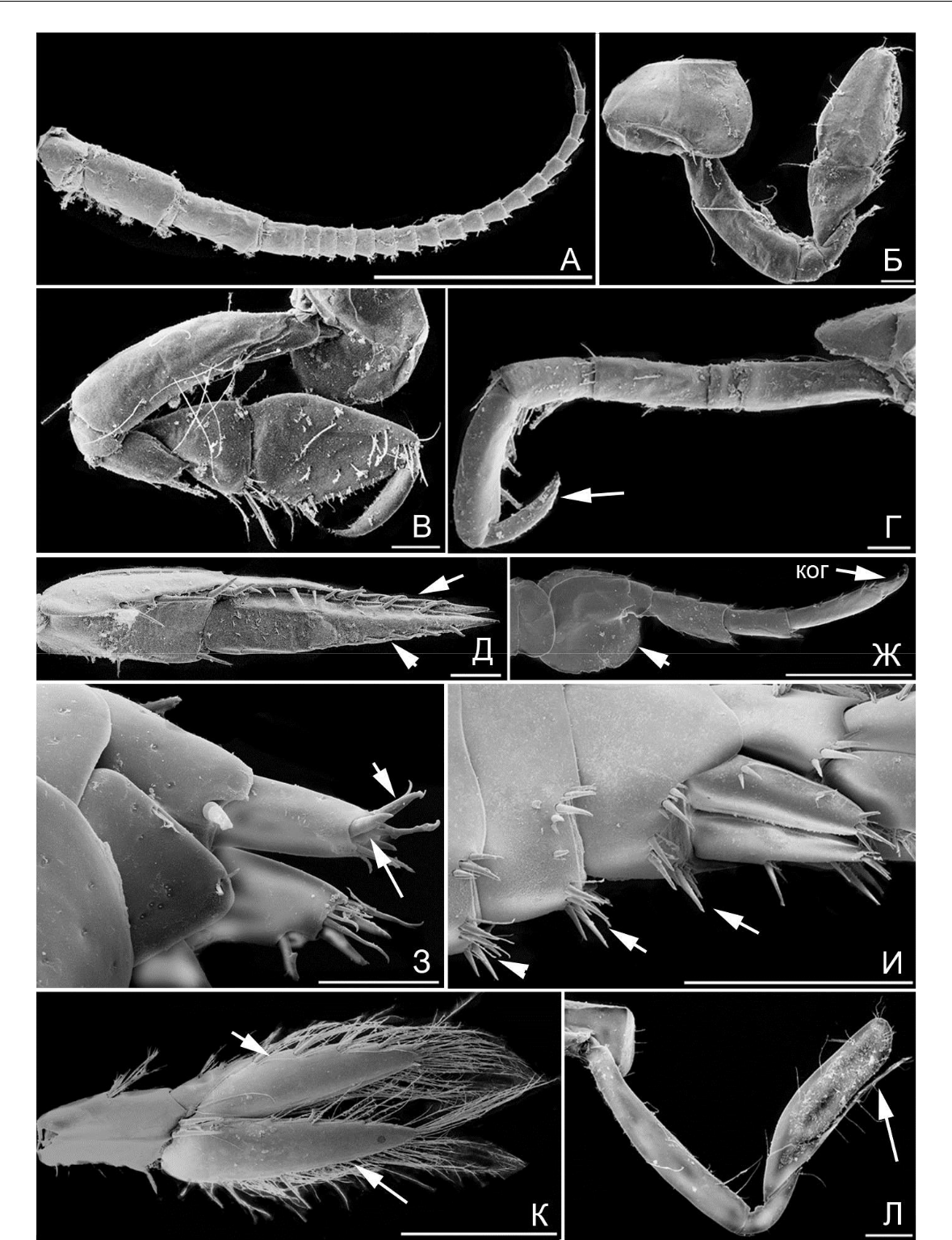
Рис. 10. Элементы морфологии амфипод
- – G. angulosus, антенна 1; Б – D. spinosa, гнатопод 1; В – D. spinosa, гнатопод 2; Г – D. spinosa,
переопод 4 (стрелкой указан коготь); Д – D. spinosa, уропод 3 (стрелками указаны выетви); Ж – A. perieri, переопод 7 (стрелкой указан базиподит, «ког» – коготь); З – A. perieri, тельсон и уропод 3 (стрелками указаны шипы ветви уропода 3); И – G. crinicornis, уросом вид латеро-дорзальный (стрелками указаны группы шипов и щетинок); К – G. aequicauda, уропод 3 (стрелками указаны ветви уропода 3); Л – P. longimanus, гнатопод 2 (стрелкой указан выступ карпуса). Масштабная линейка
0,1 мм.
48
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()

Рис. 11. Внешний вид представителей эдицеротидной (А) мегалуропиидной (Б) и талитридной жизненных форм кривофилов
- – Perioculodes longimanus (Spence Bate & Westwood, 1868); Б – Megaluropus massiliensis Ledoyer, 1976; В – Orchestia mediterranea Costa, 1853. Масштабная линейка 1 мм.
Коксальные пластинки 1–4 переона и эпимеральные пластинки 1-3 плеона образуют 2 боковых щита (рис. 11 А, функция защиты брюшка и плеопод усилена), частично отделенные зауженной частью. Карпальные членики гнатопод с выростами (рис. 10 Л, вырост указан стрелкой, функция захвата усилена). Переоподы 7-й пары чрезвычайно развиты относительно других пар, с прямым тонким и длинным когтем, не приспособленным для захвата субстрата, скорее он используется для упора на субстрат или погружения в рыхлый субстрат (рис. 11 А, смена функции захвата субстрата на функцию упора). Плеон удлиненный и массивный (рис. 11 А, функция плавания усилена), с развитыми плеоподами. Уроподы тонкие и длинные, дистально без шипов (рис. 12 А, часть ветвей указана стрелками, функция защиты уропод ослаблена за счет потери терминальных шипов).
Уроподы 3-й пары узкие, заостренные дистально (рис. 12 А, вероятно ослаблена функция рулей при плавании). Представители эдицеротидной формы обитают на рыхлых грунтах с примесью взвеси и детрита. Прямая форма когтей этой жизненной формы амфипод может затруднять их выживание в прибойной зоне, в том числе из-за ослабления или невозможности захвата за субстрат когтями во время штормов. В данную форму в Черном море входят рода Deflexilodes (2 вида), Perioculodes (1), Synchelidium (1).
Амфиподы хиалидной жизненной формы (рис. 9 Б) с хорошо развитыми коксальными пластинками 1–4 (рис. 6 Б, функция защиты брюшка с боков усилена, боковые щиты выражены) и расширенными базиподитами переопод с 5-й по 7-ю пары (рис. 10 Ж, базиподит указан стрелкой, функция защиты брюшка с боков усилена). Переоподы с развитыми загнутыми когтями и группами шипов в дистальной части проподального членика вблизи крепления когтя (рис. 10 Ж, «ког», функция захвата субстрата когтями усилена). Уроподы 3-
- пары маленькие, ветви короткие, субцилиндрические, с развитыми шипами на концах (рис. 10 З, шипы указаны стрелками, смена функции рулей на таковую упора при прыгании). Форма характерная для водорослевых сообществ, преимущественно перифитона. В Черном и частично Азовском морях к этой форме относятся рода Apohyale (3), Hyale (1), Micropythia
(1), Protohyale (2) Parhyale (2).
Амфиподы гаммаридной жизненной формы взяты за более древнюю, исходную Тело (рис. 9 В) вооружено в спинной части уросомы сочетанием шипов и щетинок или зубцов, шипов и щетинок (рис. 10 И, шипы частично указаны стрелками, выражена функция защиты от хищников). Коксальные пластинки 1–4 развитые, перекрывающиеся (рис. 9 В, функция защиты брюшка с боков усилена, боковые щиты выражены). Уроподы 3-й пары развиты, с ветвями в виде уплощенных вытянутых ланцетовидных лопастей (рис. 10 К, ветви указаны стрелками, ветви равны или одна существенно больше), снабженных различными щетинками и шипами по бокам (типичный тип рулей). Особи таксонов данной формы предпочитают скопления камней, водорослей и детрита на различных грунтах на дне. К этой форме относятся рода Amathillina (1), Chaetogammarus (3), Cheirocratus (1), Compactogammarus (1),
49
Гринцов В. А.

Рис. 12. Элементы морфологии амфипод
- – P. longimanus, уроподы (стрелками ветви); Б – M. massiliensis, коксальные пластинки 1-3; В – M. massiliensis, гнатопод 1, мерус-коготь; Г – M. massiliensis, тельсон, уроподы 3 (стрелками указаны ветви уропод); Д – O. gammarellus, антенна 1 (указана стрелкой); Ж – O. gammarellus, гнатопод 1, проподус и коготь; З – O. gammarellus, переопод 3, коготь (указан стрелкой); И – S. monoculoides, тельсон и уропод 3 (указан стрелкой). Масштабная линейка 0,1 мм.
50
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
Dikerogammarus (3), Echinogammarus (3), Euxinia (1), Gammarus (4), Gmelina (1), Gmelinopsis
(1), Iphigenella (3), Yogmelina (1), Kuzmelina (1), Melita (2), Niphargogammarus (1), Niphargoides
(1), Obessogammarus (2), Pandorites (1), Paraniphargoides (1), Pontogammarus (частично, 2 вида), Shablogammarus (1), Stenogammarus (5), Turcogammarus (1), Uroniphargoides (1).
Амфиподы мегалуропиидной жизненной формы (рис. 11 Б) с развитыми глазами, заходящими за боковые части головы (функция зрения усилена). Размеры коксальных пластинок специфически чередуются – вторая и четвертая больше, чем первая и третья (рис. 12 Б, усилена защита отдельных участков брюшной части переона). Гнатоподы обеих пар сходные морфологически, слабо развиты, без выраженного приспособления к захвату (рис. 12 В, не выражен край ладони проподуса со стороны когтя). Когти переопод вытянутые, прямые (функция захвата когтями субстрата ослаблена). Уроподы 3-й пары с развитыми листовидными широкими ветвями (рис. 12 Г, ветви указаны стрелками, по типу рулей). Представители этой формы обитают на рыхлых грунтах с примесью взвеси и детрита. Прямая форма когтей данной формы может затруднять выживание особей в прибойной зоне из-за ослабления захвата когтями за субстрат во время штормов. В мегалуропиидную форму в Черном море входят два вида из рода Megaluropus.
Амфиподы талитридной жизненной формы (рис. 11 В) с чрезвычайно маленькими и короткими антеннами 1-й пары (рис. 12 Д, антенна 1 указана стрелкой, сенсорная функция ослаблена за счет сокращения размеров и числа члеников жгутиков с чувствительными органами). Все придатки вооружены крепкими шипами, без щетинок. Все коксальные пластинки хорошо развиты (рис. 11 В, функция защиты брюшка с боков усилена). Гнатоподы 1-й пары самцов и самок маленькие, с рудиментарным краем со стороны когтя, сам коготь уменьшен в размерах (рис. 12 Ж, функция захвата предметов ослаблена). Когти переоподов мощные, загнутые (рис. 12 З, коготь указан стрелкой, функция захвата субстрата усилена). Уроподы 3-й пары маленькие, с уменьшенными короткими ветвями, близкие к конической форме, отсутствует щетка из щетинок разного типа, остаются только шипы (смена функции рулей при плавании на функцию элемента упора при прыгании). Данная форма исключительно наземная, особи обитают среди выбросов водорослей на пляжах. В Черном море в данную форму входят рода Britorchestia (1 вид), Cryptochestia (1), Orchestia (4), Platorchestia (1), Speziorchestia (1), Talitrus (1).
Амфиподы харпиниидной жизненной формы (рис. 13 А) без глаз (в Черном море), либо глаза разной формы у разных таксонов, относящихся к этой жизненной форме (преим. семейство Phoxocephalidae).

Рис. 13. Внешний вид представителей харпиниидной (А) стенотоидной (Б) и лизианассидной жизненных форм кривофилов
- – Harpinia crenulata (Boeck, 1871), по Karaman, 1993; Б – Stenothoe monoculoides (Montagu, 1815); В –
Orchomene humilis (Costa, 1853). Масштабная линейка 1 мм.
Над антеннами имеется характерный широкий козырек (функция защиты антенн усилена). Сами антенны короткие, сходные по длине (сенсорная функция ослаблена за счет сокращения числа члеников жгутиков с чувствительными органами). Переоподы 6-й пары значительно массивнее и длиннее остальных переопод; базиподиты переопод 6-й и 7-й пар разной формы. Когти переопод почти прямые (функция захвата субстрата ослаблена). Уроподы 3-й пары с ветвями, сужающимися дистально, ветви не ланцетовидные, по типу
51
Гринцов В. А.
![]()
рулей, вооружение щетинками частично редуцировано относительно гаммаридной формы. Представители данной жизненной формы обитают на илистых грунтах с примесью детрита. Прямая форма когтей харпиниидной формы может затруднять выживание особей в прибойной зоне из-за ослабления захвата за субстрат во время штормов. В Черном море харпиниидная жизненная форма представлена двумя видами рода Harpinia, обитающими а прибрежье Турции на глубине около 70 м.
Тело амфипод стенотоидной жизненной формы (рис. 13 Б), близко по контуру к овальному. Третья и особенно четвертая коксальные пластинки чрезвычайно развиты и образуют мощные боковые защитные щиты для брюшка и частично переопод (рис. 13 Б). Уроподы 3-й пары с одной палочковидной ветвью, слабо вооруженной шипами (рис. 12 И, ветвь указана стрелкой, по типу рулей). Данная форма характерна для водорослей. В Черном море отмечены представители рода Stenothoe (2 вида).
Амфиподы лизианассидной жизненной формы (рис. 13 В) со специфической формой 1-й пары антенн: стебельки вздутые, их членики бочёнковидные (рис. 14 А, членики стебелька указаны стрелкой).
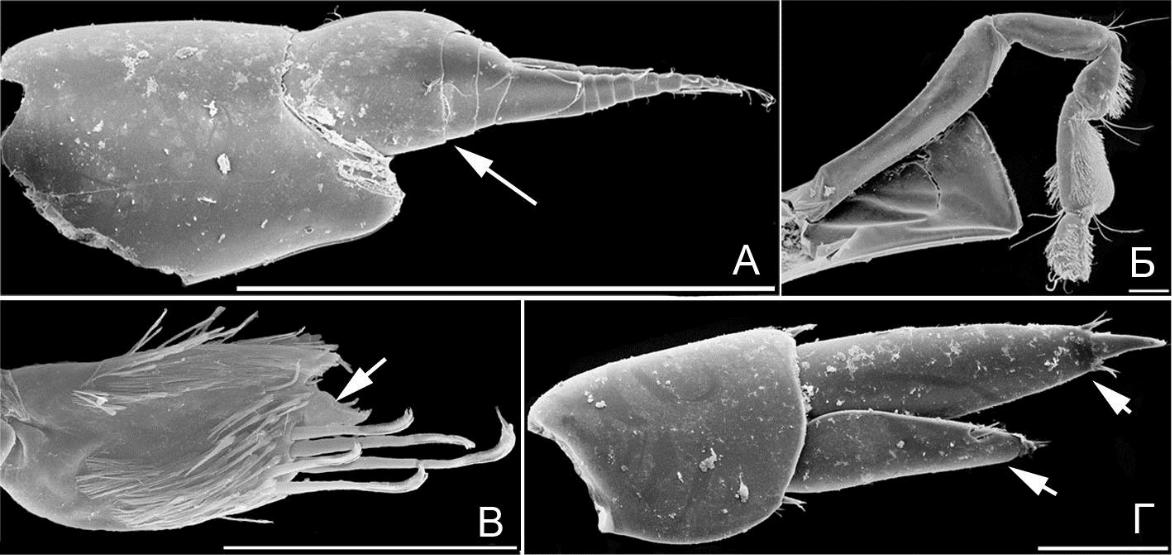
Рис. 14. Элементы морфологии амфипод
- – O. humilis, антенна 1 (стебелек указан стрелкой); Б – O. humilis, гнатопод 2; В – N. goesi reductus; гнатопод 2, проподус и коготь (коготь указан стрелкой); Г – O. humilis, уропод 3 (ветви указаны стрелкой). Масштабная линейка 0,1 мм.
Все коксальные пластинки хорошо развиты (рис. 13 В, функция бокового защитного щита усилена). Гнатоподы 2-й пары весьма специфические, тонкие; карпальный и проподальный членики близки по ширине и толщине, опушены специфически расположенной густой щеткой мелких, близких по длине щетинок (рис. 14 Б, преимущественно на задних поверхностях, вероятно выполняют роль чистящего органа), коготь маленький (рис. 14 В, коготь указан стрелкой, функция захвата предметов ослаблена). Уроподы 3-й пары с ветвями вытянуто-треугольными, без щетки щетинок (рис. 14 Г, ветви указаны стрелками, по типу рулей). У представителей данной формы нет выраженного предпочтения субстрата. В Черном море форма представлена родами Orchomene (1 вид) и Nannonyx (2).
- итоге, в результате исследований разработана общая схема выделенных экологических групп, экоморф и жизненных форм амфипод Черного и Азовского морей представленная в виде дендрограммы на основе экологии и морфологии на рисунках 15, 16.
52
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
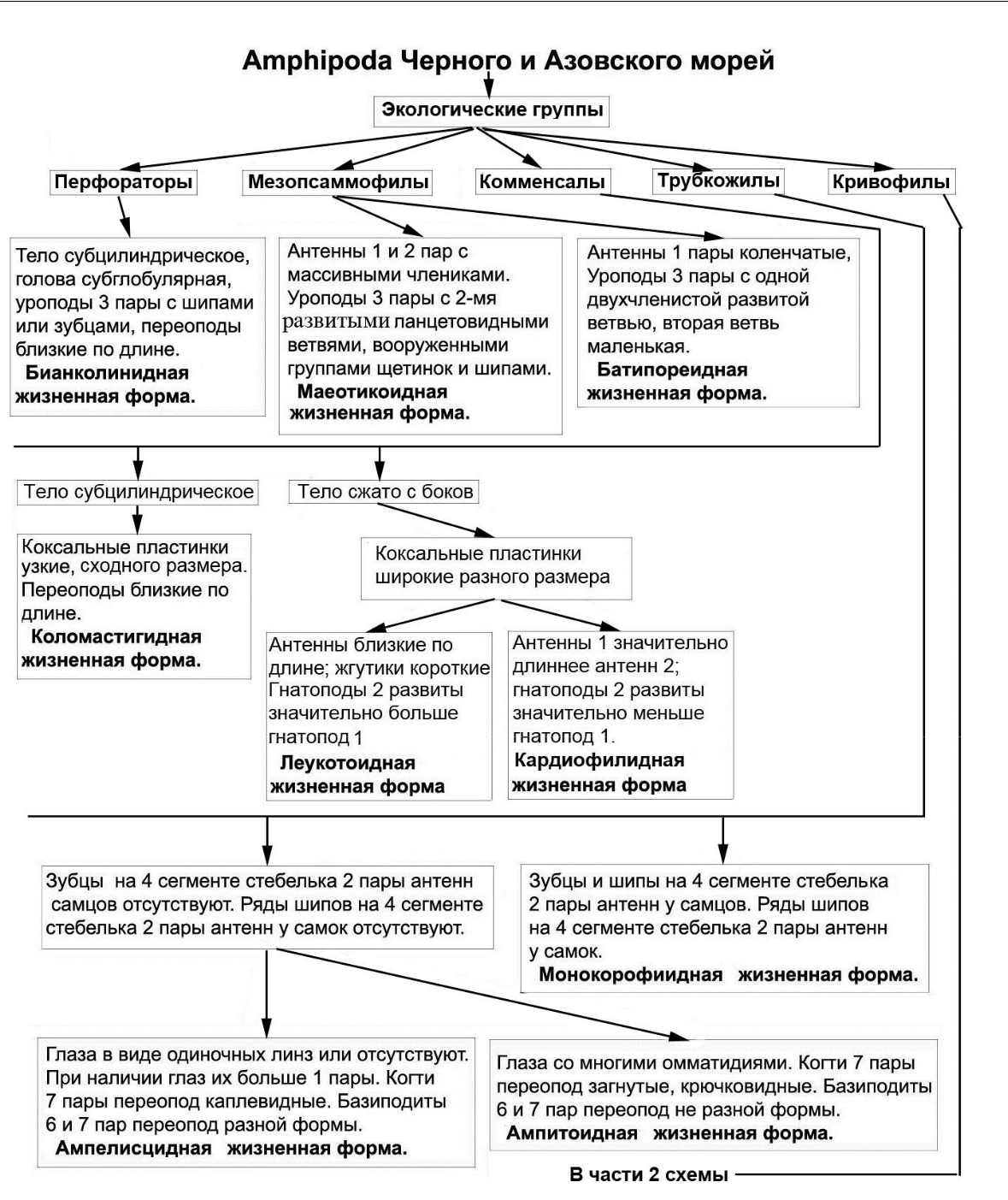
Рис. 15. Первая часть общей схемы выделения экологических групп, экоморф и жизненных форм амфипод Черного и Азовского морей
Данная схема позволяет проанализировать, какие вышеописанные идиоадаптации и другие морфологические преобразования способствовали сформированному в настоящее время эколого-морфологическому комплексу амфипод в Черном и Азовском морях. Как следует из схемы, наибольшими морфологическими преобразованиями подверглись амфиподы капреллидной экоморфы. Далее выделяется группа, способная строить трубки (трубкожилы). Выделение других групп связано со спецификой скрываться от хищников и опасностей (перфораторы, кривофилы, мезопсаммофилы, комменсалы). Образование внутри
53
Гринцов В. А.
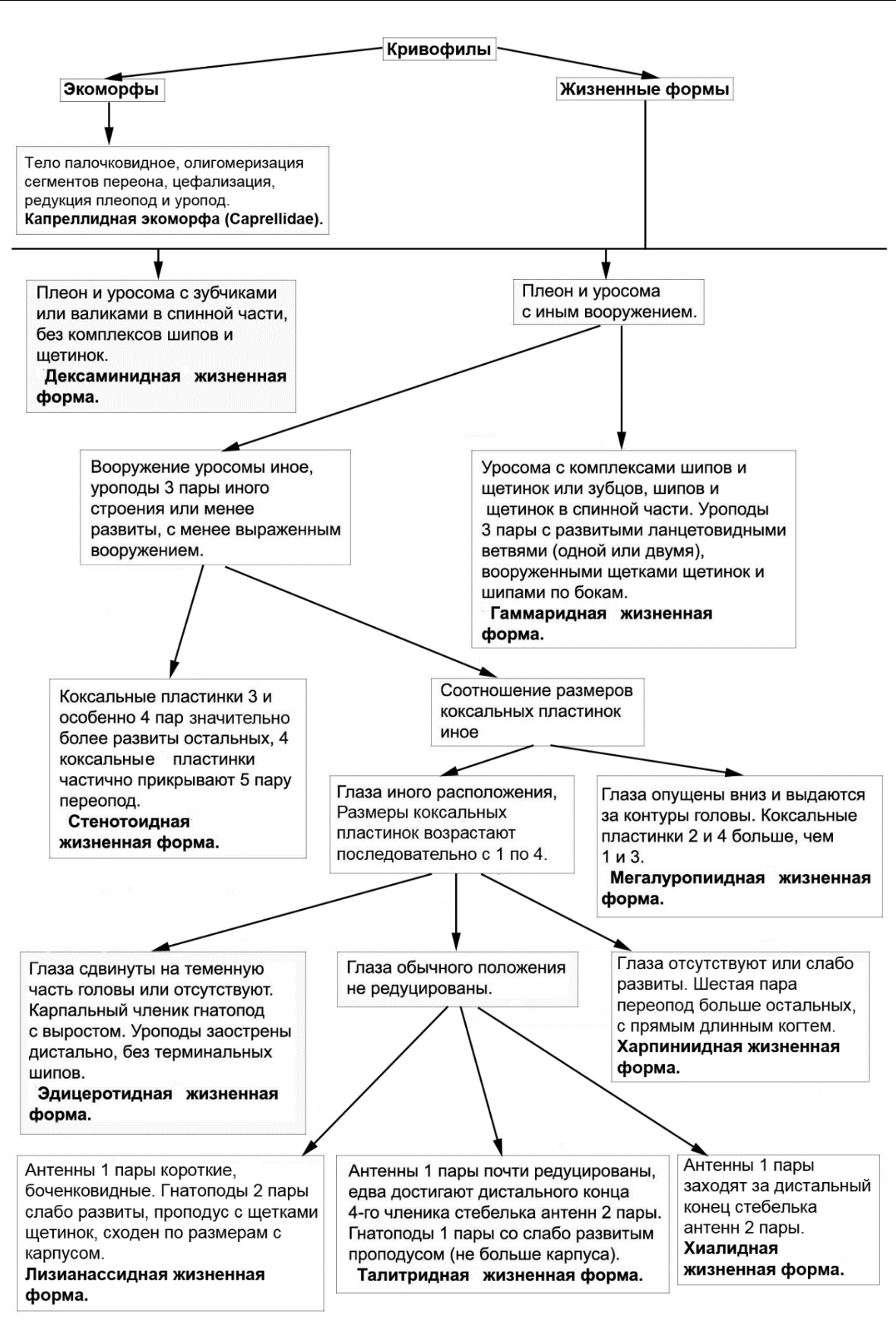
Рис. 16. Вторая часть общей схемы выделения экологических групп, экоморф и жизненных форм амфипод Черного и Азовского морей
54
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
экологических групп жизненных форм связано с появлением тех или иных идиоадаптаций, отчасти определяющих преимущественное выживание в тех или иных условиях.
Сообщества, биотопы, экотопы заселены различными экологическими группами, экоморфой и жизненными формами амфипод неравномерно. Наибольшее число жизненных форм и капреллидная экоморфа отмечены в перифитоне и водорослевых ассоциациях – монокорофиидная, ампитоидная, ампелисцидная жизненные формы трубкожилов; капреллидная экоморфа кривофилов; дексаминидная, хиалидная, гаммаридная, стенотоидная, лизианассидная жизненные формы кривофилов; бианколинидная жизненная форма перфораторов (B. algicola).
Выявленное разнообразие экологических групп и жизненных форм амфипод в данной экотопической группе (перифитон) отчасти объясняется сложной конфигурацией перифитона
- водорослевых ассоциаций, населенных множеством организмов. Так, в Черном море у берегов Крыма в перифитоне отмечено 275 видов макроорганизмов – макрофитов и беспозвоночных, относящихся к различным таксонам (Гринцов и др. 2004а, 2004б, 2005а, 2005б, 2005в; 2009). Подобное разнообразие способствует образованию множества микроусловий, обеспечивающих выживание разных амфипод и увеличивает пищевой спектр этих ракообразных. В результате к данной среде оказались приспособлены жизненные формы с разной адаптивной морфологией. Всего в перифитоне отмечено 42 вида этих ракообразных.
Следующая среда обитания относительно разнообразия экологических групп и жизненных форм – детрит и различные органические остатки на дне. В этой среде отмечены капреллидная экоморфа кривофилов; ампелисцидная жизненная форма трубкожилов;
гаммаридная, эдицеротидная, мегалуропиидная, харпиниидная, стенотоидная, лизианассидная жизненные формы кривофилов (Грезе, 1977, 1985; Киселева, 1980). Разнообразие жизненных форм и наличие капреллидной экоморфы в этой среде обитания объясняется отчасти богатой пищевой базой, включающей фрагменты растительного и частично животного происхождения. Кроме того, при достаточном слое детрита, мелкие особи различных жизненных форм могут спрятаться в нем, что способствует выживанию и как следствию увеличения разнообразия отмеченных категорий амфипод.
Несколько жизненных форм, представляющих две экологические группы, оказались связаны с различными беспозвоночными как комменсалы. Это коломастигидная, леукотоидная и кардиофилидная жизненные формы комменсалов, а также дексаминидная жизненная форма кривофилов – вид Tritaeta gibbosa (Spence Bate, 1862). В первую очередь в Черном море они связаны с губками, частично с двустворчатыми моллюсками семейства Cardiidae. Наличие нескольких жизненных форм сумевших приспособиться к комменсализму на вышеуказанных беспозвоночных, свидетельствует о возможности выбора разных путей адаптации к подобной среде.
Для остальных биотопов отмечено меньше экологических групп и жизненных форм, что свидетельствует об относительной ограниченности условий в плане выживания. Внутри древесины в ходах успешно выживает только 1 вид, относящийся к бианколинидной форме перфораторов (C. terebrans). В толще песка ниже прибойной зоны отмечен один вид мезопсаммофилов (B. guilliamsoniana). В толще песка зоны заплеска зарегистрированы 2 вида мезопсаммофилов (P. maeoticus, E. weidemanni). Толща песка в зоне супралиторали успешно заселена единственным видом талитридной формы Deshayesorchestia deshayesii (Audouin, 1826). В зоне заплеска галечно-песчанных пляжей отмечены некоторые представители гаммаридной формы кривофилов – Echinogammarus foxi Chellenberg, 1828; Echinogammarus ischnus (Stebbing, 1899); Chaetogammarus olivii (H.Milne Edwards, 1830); Echinogammarus karadagiensis Grintsov, 2009. Эти же виды, некоторые представители хиалидной жизненной формы кривофилов: Apohyale crassipes (Heller, 1866); Apohyale perieri (Lucas, 1849); Parhyale taurica Grintsov, 2009, а также один представитель ампитоидной жизненной формы – Plumulojassa ocia (Spence Bate, 1862), отмечены в зоне заплеска каменисто-валунных пляжей.
-
- сообществах бентоса по данным Киселевой (Киселева, 1980) отмечены представители ампелисцидной, ампитоидной, монокорофиидной жизненных форм трубкожилов, капреллидной экоморфы кривофилов, гаммаридной и стенотоидной жизненных форм
55
Гринцов В. А.
![]()
кривофилов. Всего в сообществах бентоса по данным (Киселева, 1980) отмечено 11 видов. Следует отметить, что почти все экологические группы, экоморфы и жизненные формы широко распространены в Мировом океане. Исключение составляют виды понто-каспийской реликтовой фауны, представляющие маеотикоидную жизненную форму маезопсаммофилов
- кардиофилидную жизненную форму комменсалов, ограниченные бассейнами Черного, Азовского и отчасти Каспийского морей. Общий список отмеченных видов в Черном и Азовском морях (с учетом литературных данных) представлен в таблице 1. Экологические группы, экоморфы и жизненные формы указаны только для видов.
Таблица 1
Список таксонов амфипод Черного и Азовского морей с указанием экологических групп,
экоморфы и жизненных форм
| Таксоны амфипод | Экологические | Экоморфы | Жизненные | |
| группы | формы | |||
| 1 | 2 | 3 | 4 |
| Ampeliscidae | |
| Ampelisca diadema (Costa, 1853) | трубкожилы |
| Ampelisca pseudosarsi Bellan-Santini | —//—//— |
| & Kaim-Malka, 1977 | |
| Ampelisca pseudospinimana Bellan- | —//—//— |
| Santini & Kaim-Malka, 1977 | |
| Ampelisca spinipes Boeck, 1861 | —//—//— |
| Ampithoidae | |
| Ampithoe ramondi Audouin, 1826 | —//—//— |
| Biancolina algicola Della Valle, 1893 | перфораторы |
| Cymadusa crassicornis (Costa, 1853) | трубкожилы |
| Pleonexes gammaroides Spence Bate, | —//—//— |
| 1856 | |
| Aoridae | |
| Microdeutopus algicola Della Valle, | —//—//— |
| 1893 | |
| Microdeutopus anomalus (Rathke, | —//—//— |
| 1843) | |
| Microdeutopus gryllotalpa Costa, | —//—//— |
| 1853 | |
| Microdeutopus stationis Della Valle, | —//—//— |
| 1893 | |
| Microdeutopus versiculatus (Spence | —//—//— |
| Bate, 1857) | |
| Atylidae | |
| Nototropis guttatus Costa, 1853 | кривофилы |
| Nototropis massiliensis (Bellan- | —//—//— |
| Santini, 1975) | |
| Bathyporeiidae | |
| Bathyporeia guilliamsoniana (Spence | мезопсаммофилы |
| Bate, 1857) | |
| Behningiellidae | |
| Cardiophilus baeri G. O. Sars, 1896 | комменсалы |
| Cardiophilus marisnigrae | —//—//— |
| Miloslawskaya, 1931 | |
| Calliopiidae | |
| Apherusa bispinosa (Spence Bate, | кривофилы |
| 1857) | |
| Apherusa chiereghinii Giordani- | —//—//— |
| Soika, 1949 |
гаммаридная
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
ампелисцидная
—//—//—
—//—//—
—//—//—
ампитоидная бианколинидная ампитоидная —//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
дексаминидная
—//—//—
батипореидная
кардиофилидная
—//—//—
дексаминидная
—//—//—
56
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
Таблица 1 (продолжение)
| 1 | 2 | 3 | 4 | |||
| Caprellidae | ||||||
| Caprella acanthifera Leach, 1814 | —// | —//— | капреллидная | —- | ||
| Caprella danilevskii Czerniavski, | —// | —//— | —// | —//— | —- | |
| 1868 | ||||||
| Caprella equilibra Say, 1818 | —// | —//— | —// | —//— | —- | |
| Caprella liparotensis Haller, 1879 | —// | —//— | —// | —//— | —- | |
| Caprella mitis Mayer, 1890 | —// | —//— | —// | —//— | —- | |
| Caprella rapax Mayer, 1890 | —// | —//— | —// | —//— | —- | |
| Phtisica marina Slabber, 1769 | —// | —//— | —// | —//— | —- | |
| Pseudoprotella phasma (Montagu, | —// | —//— | —// | —//— | —- | |
| 1804) | ||||||
| Cheirocratidae | ||||||
| Cheirocratus sundevallii (Rathke | трубкожилы | гаммаридная | ампитоидная | |||
| 1843) | ||||||
| Cheluridae | ||||||
| Chelura terebrans Philippi, 1839 | перфораторы | —// | —//— | бианколинидная | ||
| Colomastigidae | ||||||
| Colomastix pusilla Grube, 1861 | комменсалы | —// | —//— | коломастигидная | ||
| Corophiidae | ||||||
| Chelicorophium chelicorne (G. O. | трубкожилы | —// | —//— | монокорофиидная | ||
| Sars, 1895) | ||||||
| Chelicorophium curvispinum (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1895) | ||||||
| Chelicorophium maeoticum | —// | —//— | —// | —//— | —// | —//— |
| (Sowinsky, 1898) | ||||||
| Chelicorophium mucronatum (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1895) | ||||||
| Chelicorophium nobile (G. O. Sars, | —// | —//— | —// | —//— | —// | —//— |
| 1895) | ||||||
| Chelicorophium robustum (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1895) | ||||||
| Chelicorophium sowinskyi | —// | —//— | —// | —//— | —// | —//— |
| (Martynov, 1924) | ||||||
| Corophium orientale Schellenberg, | —// | —//— | —// | —//— | —// | —//— |
| 1928 | ||||||
| Corophium volutator (Pallas, 1766) | —// | —//— | —// | —//— | —// | —//— |
| Crassicorophium bonellii (H. Milne | —// | —//— | —// | —//— | —// | —//— |
| Edwards, 1830) | ||||||
| Crassicorophium crassicorne | —// | —//— | —// | —//— | —// | —//— |
| (Bruzelius, 1859); | ||||||
| Leptocheirus pilosus Zaddach, 1844 | —// | —//— | —// | —//— | ампитоидная | |
| Medicorophium runcicorne (Della | —// | —//— | —// | —//— | монокорофиидная | |
| Valle, 1893) | ||||||
| Monocorophium acherusicum (Costa, | —// | —//— | —// | —//— | —// | —//— |
| 1853) | ||||||
| Monocorophium insidiosum | —// | —//— | —// | —//— | —// | —//— |
| (Crawford, 1937) | ||||||
| Dexaminidae | ||||||
| Dexamine spiniventris (Costa, 1853) | кривофилы | —// | —//— | дексаминидная | ||
| D. spinosa (Montagu, 1813) | —// | —//— | —// | —//— | —// | —//— |
| D. thea Boeck, 1861 | —// | —//— | —// | —//— | —// | —//— |
| Tritaeta gibbosa (Spence Bate, 1862) | —// | —//— | —// | —//— | —// | —//— |
57
Гринцов В. А.
![]()
Таблица 1 (продолжение)
| 1 | 2 | 3 | 4 | |||
| Gammarellidae | ||||||
| Gammarellus angulosus (Rathke, | —// | —//— | —// | —//— | —// | —//— |
| 1843) | ||||||
| Gammarellus carinatus (Rathke, | —// | —//— | —// | —//— | —// | —//— |
| 1837) | ||||||
| Gammaridae | ||||||
| Amathillina cristata (G. O. Sars, | ||||||
| 1894) | —// | —//— | —// | —//— | гаммаридная | |
| Chaetogammarus placidus (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1896) | ||||||
| Chaetogammarus olivii (H. Milne | —// | —//— | —// | —//— | —// | —//— |
| Edwards, 1830) | ||||||
| Dikerogammarus villosus | ||||||
| (Sowinskyi, 1894) | —// | —//— | —// | —//— | —// | —//— |
| Dikerogammarus haemobaphes | —// | —//— | —// | —//— | —// | —//— |
| (Eichwald, 1841) | ||||||
| Dikerogammarus gruberi Mateus & | —// | —//— | —// | —//— | —// | —//— |
| Mateus, 1990 | ||||||
| Dikerogammarus istanbulensis | —// | —//— | —// | —//— | —// | —//— |
| Özbek, 2011 | ||||||
| Gmelina costata G. O. Sars, 1894 | —// | —//— | —// | —//— | —// | —//— |
| Gmelinopsis tuberculata G. O. Sars, | —// | —//— | —// | —//— | —// | —//— |
| 1896 | ||||||
| Yogmelina pusilla (G. O. Sars, 1896) | —// | —//— | —// | —//— | —// | —//— |
| Kuzmelina kusnezowi (Sowinskyi, | —// | —//— | —// | —//— | —// | —//— |
| 1894) | ||||||
| Shablogammarus subnudus (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 18961896) | ||||||
| Echinogammarus foxi (Schellenberg, | —// | —//— | —// | —//— | —// | —//— |
| 1928) | ||||||
| Echinogammarus ischnus (Stebbing, | —// | —//— | —// | —//— | —// | —//— |
| 1899) | ||||||
| Echinogammarus karadagiensis | —// | —//— | —// | —//— | —// | —//— |
| Grintsov, 2009 | ||||||
| Echinogammarus warpachowsky (G. | —// | —//— | —// | —//— | —// | —//— |
| O. Sars, 1894) | ||||||
| Gammarus aeqiucauda (Martynov, | —// | —//— | —// | —//— | —// | —//— |
| 1931) | ||||||
| Gammarus crinicornis Stock, 1966 | —// | —//— | —// | —//— | —// | —//— |
| Gammarus insensibilis Stock, 1966 | —// | —//— | —// | —//— | —// | —//— |
| Gammarus subtypicus Stock, 1966 | —// | —//— | —// | —//— | —// | —//— |
| Hyalidae | ||||||
| Apohyale crassipes (Heller, 1866) | —// | —//— | —// | —//— | хиалидная | |
| Apohyale perieri (Lucas, 1849) | —// | —//— | —// | —//— | —// | —//— |
| Apohyale prevostii (H. Milne | —// | —//— | —// | —//— | —// | —//— |
| Edwards, 1830) | ||||||
| Hyale pontica Rathke, 1836 | —// | —//— | —// | —//— | —// | —//— |
| Protohyale (Boreohyale) camptonyx | —// | —//— | —// | —//— | —// | —//— |
| (Heller, 1866) | ||||||
| Protohyale (Protohyale) schmidti | —// | —//— | —// | —//— | —// | —//— |
| (Heller, 1866) | ||||||
| Parhyale aquilina (Costa, 1857) | —// | —//— | —// | —//— | —// | —//— |
| Parhyale taurica Grintsov, 2009 | —// | —//— | —// | —//— | —// | —//— |
58
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
Таблица 1 (продолжение)
| 1 | 2 | 3 | 4 | |||
| Iphigenellidae | ||||||
| Iphigenella acanthopoda G. O. Sars, | —// | —//— | —//— | //— | гаммаридная | |
| 1896 | ||||||
| Iphigenella andrussowi G. O. Sars, | —// | —//— | —//— | //— | —// | —//— |
| 1894 | ||||||
| Iphigenella shablensis Carausu, 1943 | —// | —//— | —//— | //— | —// | —//— |
| Ischyroceridae | ||||||
| Centraloecetes dellavallei (Stebbing, | трубкожилы | —//— | //— | ампитоидная | ||
| 1899) | ||||||
| Ericthonius difformis H. Milne | —// | —//— | —//— | //— | —// | —//— |
| Edwards, 1830 | ||||||
| Ericthonius punctatus (Spence Bate, | —// | —//— | —//— | //— | —// | —//— |
| 1857) | ||||||
| Ericthonius rubricornis (Simpson, | —// | —//— | —//— | //— | —// | —//— |
| 1853) | ||||||
| Jassa marmorata Holmes, 1905 | —// | —//— | —//— | //— | —// | —//— |
| Jassa ocia (Spence Bate, 1862) | —// | —//— | —//— | //— | —// | —//— |
| Jassa pusilla (G.O. Sars, 1894) | —// | —//— | —//— | //— | —// | —//— |
| Kuriidae | ||||||
| Micropythia carinata (Spence Bate, | кривофилы | —//— | //— | хиалидная | ||
| 1862) | ||||||
| Leucothoidae | ||||||
| Leucothoe spinicarpa (Abildgaard, | комменсалы | —//— | //— | леукотоидная | ||
| 1789) | ||||||
| Lysianassidae | ||||||
| Nannonyx propinguus Chevreux 1911 | кривофилы | —//— | //— | лизианассидная | ||
| Nannonyx goesi reductus Greze, 1975 | —//— | //— | ||||
| Megaluropiidae | ||||||
| Megaluropus agilis Hoek, 1889 | кривофилы | —//— | //— | мегалуропиидная | ||
| Megaluropus massiliensis Ledoyer, | —// | —//— | —//— | //— | —// | —//— |
| 1976 | ||||||
| Melitidae | ||||||
| Melita palmata (Montagu, 1804) | —// | —//— | —//— | //— | гаммаридная | |
| Microprotopidae | ||||||
| Microprotopus longimanus Chevreux, | трубкожилы | —//— | //— | ампитоидная | ||
| 1887 | ||||||
| Microprotopus maculatus Norman, | —// | —//— | —//— | //— | —// | —//— |
| 1867 | ||||||
| Oedicerotidae | ||||||
| Deflexilodes gibbosus (Chevreux, | кривофилы | —//— | //— | эдицеротидная | ||
| 1888) | ||||||
| Deflexilodes griseus (Della Valle, | —// | —//— | —//— | //— | —// | —//— |
| 1893) | ||||||
| Perioculodes longimanus (Spence | —// | —//— | —//— | //— | —// | —//— |
| Bate& Westeood, 1868) | ||||||
| Synchelidium maculatum Stebbing, | —// | —//— | —//— | //— | —// | —//— |
| 1906 | ||||||
| Phoxocephalidae | ||||||
| Harpinia crenulata (Boeck, 1871) | —// | —//— | —//— | //— | харпиниидная | |
| H. dellavallei Chevreux, 1910 | —// | —//— | —//— | //— | —// | —//— |
| Photidae | ||||||
| Megamphopus cornutus Norman, | трубкожилы | —//— | //— | ампитоидная | ||
| 1869 | ||||||
| Photis longicaudata (Spence Bate & | —// | —//— | —//— | //— | —// | —//— |
| Westwood, 1862) | ||||||
59
Гринцов В. А.
![]()
Таблица 1 (продолжение)
| 1 | 2 | 3 | 4 | |||
| Pontogammaridae | ||||||
| Compactogammarus compactus (G. | кривофилы | —// | —//— | гаммаридная | ||
| O. Sars, 1895) | ||||||
| Euxinia sarsi (Sowinsky, 1898) | —// | —//— | —// | —//— | —// | —//— |
| Euxinia weidemanni (G. O. Sars, | мезопсаммофилы | —// | —//— | маеотикоидная | ||
| 1896) | ||||||
| Niphargogammarus intermedius | кривофилы | —// | —//— | гаммаридная | ||
| (Carausu, 1943) | ||||||
| Niphargoides corpulentus G. O. Sars, | —// | —//— | —// | —//— | —// | —//— |
| 1895 | ||||||
| Obesogammaruss crassus (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1894) | ||||||
| Obesogammaruss obesus (G. O. Sars, | —// | —//— | —// | —//— | —// | —//— |
| 1894) | ||||||
| Pandorites podoceroides G. O. Sars, | —// | —//— | —// | —//— | —// | —//— |
| 1895 | ||||||
| Paraniphargoides motasi (Carausu, | —// | —//— | —// | —//— | —// | —//— |
| 1943) | ||||||
| Pontogammarus abbreviatus (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1894) | ||||||
| Pontogammarus aestuarius | —// | —//— | —// | —//— | —// | —//— |
| (Derzhavin, 1924) | ||||||
| Pontogammarus maeoticus | мезопсаммофилы | —// | —//— | маеотикоидная | ||
| (Sovinskij, 1894) | ||||||
| Pontogammarus robustoides (G. O. | кривофилы | —// | —//— | гаммаридная | ||
| Sars, 1894) | ||||||
| Stenogammarus compressus (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1894) | ||||||
| Stenogammarus deminutus (Stebbing, | —// | —//— | —// | —//— | —// | —//— |
| 1906) | ||||||
| Stenogammarus kereuschi Derzhavin | —// | —//— | —// | —//— | —// | —//— |
| & Pjatakova, 1962 | ||||||
| Stenogammarus (Stenogammarus) | —// | —//— | —// | —//— | —// | —//— |
| macrurus (G. O. Sars, 1894) | ||||||
| Stenogammarus similis (G. O. Sars, | —// | —//— | —// | —//— | —// | —//— |
| 1894) | ||||||
| Turcogammarus aralensis (Uljanin, | —// | —//— | —// | —//— | —// | —//— |
| 1875) | ||||||
| Turcogammarus turcarum (Stock, | —// | —//— | —// | —//— | —// | —//— |
| 1974) | ||||||
| Uroniphargoides spinicaudatus | —// | —//— | —// | —//— | —// | —//— |
| (Carausu, 1943) | ||||||
| Stenothoidae | ||||||
| Stenothoe marina (Spence Bate, | —// | —//— | —// | —//— | стенотоидная | |
| 1856) | ||||||
| Stenothoe monoculoides (Montagu, | —// | —//— | —// | —//— | —// | —//— |
| 1813) | ||||||
| Talitridae | ||||||
| Britorchestia brito (Stebbing, 1891) | —// | —//— | —// | —//— | талитридная | |
| Cryptorchestia cavimana (Heller, | —// | —//— | —// | —//— | —// | —//— |
| 1865) | ||||||
| Deshayesorchestia deshayesii | —// | —//— | —// | —//— | —// | —//— |
| (Audouin, 1826) | ||||||
| Orchestia bottae H. Milne Edwards, | —// | —//— | —// | —//— | —// | —//— |
| 1840 | ||||||
60
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
Таблица 1 (продолжение)
| 1 | 2 | 3 | 4 | |||
| Orchestia gammarellus (Pallas, 1766) | —// | —//— | —// | —//— | —// | —//— |
| Orchestia mediterranea Costa, 1853 | —// | —//— | —// | —//— | —// | —//— |
| Orchestia montagui Audouin, 1826 | —// | —//— | —// | —//— | —// | —//— |
| Platorchestia platensis (Krøyer, | —// | —//— | —// | —//— | —// | —//— |
| 1845) | ||||||
| Speziorchestia stephenseni Cecchini, | —// | —//— | —// | —//— | —// | —//— |
| 1928 | ||||||
| Talitrus saltator (Montagu, 1808) | —// | —//— | —// | —//— | —// | —//— |
| Tryphosidae | ||||||
| Orchomene humilis (Costa, 1853) | —// | —//— | —// | —//— | лизианассидная | |
ВЫВОДЫ
-
- Черном и Азовском морях отмечено 5 экологических групп амфипод – трубкожилы, кривофилы, комменсалы, мезопсаммофилы, перфораторы. У трубкожилов выделены монокорофиидная, ампитоидная и ампелисцидная жизненные формы, идиоадаптации которых затронули следующие морфологические признаки: характер уплощения тела (с боков или сверху вниз), когти переопод, антенны 2-й пары, коксальные пластинки, переоподы, уроподы третьей пары. Сочетание данных идиоадаптаций специфично для каждой жизненной формы.
У кривофилов Черного и Азовского морей отмечена капреллидная экоморфа (семейство Caprellidae), отличающаяся палочковидным телом, олигомеризацией сегментов тела, цефализацией, редукцией придатков – переопод, плеопод, уропод, усилением проподальных члеников и когтей переопод.
Отмечено 9 жизненных форм кривофилов: дексаминидная, хиалидная, гаммаридная,
эдицеротидная, мегалуропиидная, талитридная, харпиниидная, стенотоидная, лизианассидная идиоадаптации которых затронули следующие морфологические элементы: рострум, глаза, вооружение спины, антенны 1 и 2 пар, коксальные пластинки, гнатоподы 1-й
- 2-й пар; уроподы, особенно 3-й пары, переоподы, когти переопод, вооружение уросомы. Сочетание морфологических идиоадаптаций специфично для каждой жизненной формы.
Бианколинидная жизненная форма перфораторов отличается цилиндрилизацией тела, полушарообразной головой, слабой развитостью коксальных пластинок, близкой длиной переопод. Идиоадаптации батипореидной и маеотикоидной жизненных форм мезопсаммофилов затронули антенны, гнатоподы, переоподы, уроподы 3-й пары. У коломастигидной, леукотоидной и кардиофилидной жизненных форм комменсалов идиоадаптации затронули антенны, коксальные пластинки, характер уплощенности тела, гнатоподы, переоподы, уроподы 3-й пары. Все вышеуказанные жизненные формы амфипод кроме кардиофилидной и маеотикоидной широко распространены в Мировом океане.
Наибольшее разнообразие экологических групп и жизненных форм в Черном море отмечено для перифитона и скоплений органических остатков (включая макрофиты) на дне. Меньше заселяется древесина, оказавшаяся в морской воде; заплеск галечно-песчанного пляжа; толща песка в зоне супралиторали пляжей.
Исследование выполнено в рамках темы госзадания «Исследование механизмов управления продукционными процессами в биотехнологических комплексах с целью разработки научных основ получения биологически активных веществ и технических продуктов морского генезиса» (№ гос. регистрации 121030300149-0)
61
Гринцов В. А.
![]()
Список литературы
Алеев Ю. Г. Жизненная форма как система адаптаций // Успехи современной биологии. – 1980. – Т. 90, вып.
- – С. 462–477.
Булычева А. И. Морские блохи морей СССР и сопредельных вод. – М-Л.: Изд-во АН СССР, 1957. – 185 с.
Василенко С. В. Капреллиды (морские козочки) морей СССР и сопредельных вод. – Л.: Наука, 1974. – 287 с.
Дедю И. И. Экологический энциклопедический словарь. – Кишинев.: Главная редакция Молдавской советской энциклопедии, 1990. – 406 с.
Димо Н. А., Келлер Б. А. В области полупустыни. – Саратов.: Изд. Саратовского губернского земства.
Почвенная. лаборатория, 1907. – 608 с.
Грезе И. И. Амфиподы Чёрного моря и их биология. – Киев.: Наук. думка, 1977. – 154 с.
Грезе И. И. Фауна Украины. Высшие ракообразные. Амфиподы. – Киев.: Наук. думка, 1985. – Т. 26, вып. 5. – 172 с.
Гринцов В. А., Мурина В. В., Евстегнеева И. К., Макаров М. А Сообщество обрастания на искусственном рифе в п. Курортное (Карадаг) // Карадаг. Гидробиологические исследования (сб. науч. тр. посвящ. 90-летию Карадаг науч. станции им. Т. И. Вяземского и 25-летию Карадаг. природ. Заповедника НАНУ). – Симферополь: 2004а. – Т. 2. – С. 152–165.
Гринцов В. А. Отряд амфиподы или разноногие раки (амфиподы) // Карадаг. Гидробиологические исследования (сб. науч. тр. посвящ. 90-летию Карадаг науч. станции им. Т. И. Вяземского и 25-летию Карадаг. природ. Заповедника НАНУ). – Симферополь, 2004б. – Т. 2. – С. 391–397.
Гринцов В. А., Костенко Н. С., Мурина В. В., Евстигнеева И.К. Биоразнообразие и структура сообщества акватории Карадагского природного заповедника // Заповедники Крыма: заповедное дело, биоразнообразие, экообразование: Матер. III науч. конф. (Симферополь, 22 апреля 2005.г.). – Симферополь, 2005а. – Часть 2. Зоология беспозвоночных, Зоология позвоночных. Экология. – С. 207–212.
Гринцов В. А., Мурина В. В., Евстигнеева И. К. Биоразнообразие и структура сообщества обрастания твердых субстратов Карадагского природного заповедника (Чёрное море) // Морской экологический журнал – 2005б. – Т. 4, № 3. – С. 37–47.
Гринцов В. А., Мурина В. В., Евстигнеева И. К. Новые сведения о биоразнообразии сообщества обрастаний твердых субстратов в прибрежной зоне Крыма // Науковi записки Тернопiльського нацiонального педагогiчного. унiверситету Серiя. Біологія. Спецiальний випуск: Гідроекологія – 2005в. – № 4 (27). – С. 54–55.
Гринцов В. А. Биоразнообразие и экология бокоплавов прибрежья Карадага // Карадаг – 2009 (сб. науч. Тр. посвящ. 95-летию Карадаг науч. станции и 30-летию Карадаг. природ. Заповедника НАНУ). – Севастополь, 2009б. – С. 361–365.
Гринцов В. А. Морфофункциональные параметры плавательных ножек плеопод (Амфипод) и некоторые моменты филогении и систематики // Бюллетень МОИП – 2010. – Т. 115, вып. 5. – С. 36–42.
Гринцов В. А. Морфологические различия видов рода Hyale (Amphipoda, Hyalidae) из Чёрного моря (прибрежье Крыма) // Весник Зоологии. – 2011а. – № 45 (5). – С. 447–455.
Гринцов В. А. Морфологические различия самок рода Orchestia и Platorchestia (Amphipoda, Talitridae) супралиторали Чёрного моря (Крым) // Зоологический журнал – 2011б. – Т. 90, № 2. – С. 143–148.
Гринцов В. А. Динамика структуры популяции двух видов бокоплавов рода Echinogammarus (Gammaridae, Amphipoda) из бухты Ласпи (Крым, Чёрное море) // Морской биологический журнал – 2016в. – Т. 1, № 3. – С. 22– 26.
Гринцов В. А. Биоразнообразие и динамика численности бокоплавов на коллекторах мидийной фермы (Севастополь, Черное море) // Морской биологический журнал – 2017. – Т 2, № 4. – С. 30–37.
Гринцов В. А. Амфиподы морских ферм прибрежья Севастополя (Черное море) // Водные биоресурсы и среда обитания – 2021. – Т. 4, № 4. – С. 27–35.
Гринцов В. А., Бондаренко Л. В., Тимофеев В. А. Новый вид амфиподы Melita Leach, 1814 (Crustacea, Amphipoda, Melitidae) для Азово-Черноморского бассейна // Российский журнал биологических инвазий – 2022а. –
- 5, № 1. – С. 41–54. https://doi.org/10.35885/1996-1499-15-1-41-54
Гринцов В. А. Амфиподы Черного моря: иллюстрированный атлас-определитель // ФИЦ «Институт биологии
южных морей им. А.О. Ковалевского РАН». – Севастополь: ФИЦ ИНБЮМ, 2022б. – 476 c.
Гурьянова Е. Ф. Амфиподы морей СССР. – Л.: Изд-во АН СССР, 1951. – 1029 с.
Киселева М. И. Бентос рыхлых грунтов Чёрного моря. – Киев, Наук. думка, 1981. – 168 с.
Маккавеева Е. Б. Беспозвоночные зарослей макрофитов Чёрного моря. – Киев, Наукова думка,. 1979. – 228 с. Мордухай-Болтовской Ф. Д., Грезе И. И., Василенко С. В. Отряд амфиподы или разноногие. Amphipoda // –
В кн.: Определитель фауны Чёрного и Азовского морей. – Киев, Наук. думка, 1969. – Т. 2. – С. 440–494.
Северцов А. Н. Морфологические закономерности эволюции. – М-Л.: Изд-во. АН СССР, 1949. – Т. 5. – С. 1–
536.
Формозов А. Н. Млекопитающие Северной Монголии по сборам экспедиции 1926 г. – М.: 1929. – 144 с. Arfianti T., Wilson S., Costello M. J. Progress in the discovery of amphipod crustaceans. – 2018. PeerJ 6:e5187; DOI
10.7717/peerj.5187
Barnard J. L., Karaman G. S. The families and genera of marine Gammaridean Amphipoda (except marine Gammaroids) // Records of the Australian Museum. – 1991. – Suppl. 13 (Parts 1–2). – P. 866.
62
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
Grintsov V. A new amphipod species Echinogammarus karadagiensis sp. n. (Amphipoda, Gammaridae) from Crimean coasts (Black Sea, Ukraine) // Vestnik Zoologii. – 2009. – Vol. 43, N 2. – P. 23–26.
Grintsov V. On finding Dexamine thea (Amphipoda, Dexaminidae) in the Ukrainian territorial waters (Crimea, Black Sea) // Vestnik zoologii – 2010. – Vol. 44, N 3. – P. 281–283.
Grintsov V. Dynamyx of population structure of the Amphipod Dexamine thea (Dexaminidae, Amphipoda), a new species for the Black Sea in the coastal areas in Laspi Bay (Black Sea, Sevastopol) // Russian Journal of Biological Invasions. – 2014. – Vol. 5, N 1. – Р. 18–20.
Grintsov V. On finding of Monocorophium insidiosum Crawford, 1937 (Amphipoda, Corophiidae) in the coastal waters of Crimea (Black sea), a new species for this region // Marine biological journal. – 2018. – Vol. 3, N 2. – P. 33–39.
Grintsov V. А. First finding of Centraloecetes cf. neapolitanus (Schiecke, 1978) (Ischyroceridae, Amphipoda) in coastal zone of Sevastopol (Crimea, Black Sea) // Труды Карадагской научной станции им. Т. И. Вяземского – природного заповедника РАН. – 2021а. – № 2 (18). – С. 3–11.
Grintsov V. A. First finding of Caprella сf. equilibra Say, 1818 (Amphipoda, Capreliidae) in coastal zone south-west of Crimea (Black Sea) // Труды Карадагской научной станции им. Т. И. Вяземского – природного заповедника РАН. – 2021б. – № 4 (20). – С. 3–9.
Grintsov V., Sezgin M. Manual for identification of Amphipoda from The Black Sea // Sevastopol. Digit Print. – 2011. – 151 p.
Kudrenko S. A. Amphipod (Crustacea, Amphipoda) communities in the north-western part of the Black Sea // Vestnik zoologii – 2016. – Vol. 50 (5). – P. 387–394.
Petrescu I. Contribution to the knowledge of Amphipods (Crustacea: Amphipoda) from Romania. 7. Amphipods from Agigea (Black Sea) // Travaux du Museum d’Historie Naturelle “Grigore Antipa”. – 1998. – Vol. 15. – P. 51–73.
Sezgin M., Kocataş A., Katağan T., Amphipod Fauna of the Turkish Central Black Sea Region. // Turkish Journal of Zoology – 2001. – Vol. 25. – P. 57–61.
Uzunova S. Checklist of marine Amphipoda (Crustacea, Malacostraca) from the Bulgarian Black Sea area // Известия на съюза на учените – Варна, Серия «Морски науки». – 2012. – С. 72–79.
Grintsov V. A. Ecological groups, ecomorphs and life forms of amphipods (Crustacea, Amphipoda) of the Black Sea and the Sea of Azov // Ekosistemy. 2023. Iss. 33. P. 38–63.
On the basis of the analysis of morphology, biology, ethology, ecology of 141 species of amphipods of the Black Sea and the Sea of Azov, five ecological groups were identified: tubebuilders, perforators, commensals, kryvophiles (from the Greek «kryvo» – to hide, «phileo» – to love) and mesopsammophiles (from the Greek «meso» – inside, «psammo» – sand, «phileo» – to love). Moreover, ecomorphs and life forms are defined within these five ecological groups. The caprellid ecomorph (family Caprellidae) and nine life forms named after the taxa included in them are designated in the kryvophiles: hialid (genus Hyale), dexaminid (genus Dexamine), gammarid (genus Gammarus), edicerotid (family Oedicerotidae), megaluropiid (genus Megaluropus), talitrid (family Talitridae), harpiniid (genus Harpinia), stenotoid (genus Stenothoe), Lisianassid (family Lisianassidae). The life forms of all five ecological groups are not restricted to the taxa they are named after. Tubebuilders are divided into ampeliscid (genus Ampelisca), ampitoid (genus Ampithoe) and monocorophiid (genus Monocorophium) life forms. The perforators have one life form – biancolinid (genus Biancolina). The commensals have three life forms: cardiophilid (genus Cardiophilus), colomastigid (genus Colomastix) and leukotoid (genus Leucothoe). Two life forms were identified in mesopsammophiles – bathyporeid (genus Bathyporeia) and maeoticoid (species Pontogammarus maeoticus). This article analyzes ecological groups, ecomorphs and life forms, as well as their distribution in communities, biotopes, habitats or ecotopes in accordance with morphological idioadaptations of amphipods, ethology, ecology and biology.
Key words: Amphipoda, ecomorphs, ecological groups, live forms, Black Sea, Sea of Azov.
Поступила в редакцию 11.01.23
Принята к печати 05.02.23
63
Институт биологии южных морей имени А. О. Ковалевского РАН Севастополь, Россия
VGrintsov@gmail.com
На основе анализа морфологии, биологии, этологии и экологии 141 вида амфипод Черного и Азовского морей выделено пять экологических групп: трубкожилы, перфораторы, комменсалы, кривофилы (от греческого «kryvo»
– прятаться, «phileо» – любить) и мезопсаммофилы (от греческого «meso» – внутри, «psammo» – песок, «phileo» – любить). В пределах данных пяти экологических групп обозначены экоморфы и жизненные формы. У кривофилов выделена капреллидная экоморфа (семейство Caprellidae) и 9 жизненных форм, названных по таксонам, входящим
- них: хиалидная (род Hyale), дексаминидная (род Dexamine), гаммаридная (род Gammarus), эдицеротидная (семейство Oedicerotidae), мегалуропиидная (род Megaluropus), талитридная (семейство Talitridae), харпиниидная (род Harpinia), стенотоидная (род Stenothoe), лизианассидная (семейство Lisianassidae). Жизненные формы всех 5 экологических групп не ограничены таксонами, по которым они названы. Трубкожилы разделены на ампелисцидную (род Ampelisca), ампитоидную (род Ampithoe) и монокорофиидную (род Monocorophium) жизненный формы. У перфораторов выделена одна жизненная форма – бианколинидная (род Biancolina). У комменсалов обозначены три жизненных формы: кардиофилидная (род Cardiophilus), коломастигидная (род Colomastix) и леукотоидная (род Leucothoe). У мезопсаммофилов выделены две жизненных формы – батипореидная (род Bathyporeia) и маеотикоидная – вид Pontogammarus maeoticus (Sovinskij, 1894). В настоящей статье анализируется экологические группы, экоморфы и жизненные формы, а также их распределение по сообществам, биотопам, средам обитания или экотопам в соответствие с морфологическими идиоадаптациями амфипод, а также этологией, экологией и биологией этих ракообразных.
Ключевые слова: Amphipoda, экоморфы, экологические группы, жизненные формы, Черное море, Азовское
море.
ВВЕДЕНИЕ
Экологические группы (Димо, Келлер, 1907), экоморфы (Алеев, 1980) и жизненные формы (термин введен А. Гумбольтом в 1806 году, в зоологии одним из первых в современном значении приведен А. Н. Формозовым (1929)) живых организмов формируются
- процессе эволюции под влиянием той или иной среды обитания, в том числе через идиоадаптации, определяющие успех выживания организмов в предпочитаемых экотопах, биотопах, сообществах. Идиоадаптации не приводят к общему повышению уровня организации организмов как ароморфозы, но преобразуют частные изменения функции органов (Северцов, 1949).
- данной работе понятие «экологическая группа» описывается следующей формулировкой: «экологи́ческая гру́ппа — совокупность видов, характеризующаяся сходными потребностями в величине какого-либо экологического фактора и возникшими в результате его воздействия в процессе эволюции сходными анатомо-морфологическими и иными признаками, закрепившимися в генотипе». Для описания экоморф взята формулировка: «экоморфа – целостная система взаимообусловленных эколого-морфологических адаптаций, определяющая общую конструкцию тела организма в соответствие с конкретным направлением эволюции вида в условиях конкретного биотопа» (Алеев, 1980). Для описания жизненных форм использовали формулировку: «жизненная форма – внешний облик растений или животных, как систематически близких, так и далеких, отражающий их приспособленность к условиям среды» (Дедю, 1990).
ISSN 2414-4738 Published by V. I. Vernadsky Crimean Federal University, Simferopol
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
Цель данных исследований – проанализировать морфологические идиоадаптации, определяющие экологические группы, экоморфы и жизненные формы амфипод Черного и Азовского морей, а также распределение этих категорий по сообществам, биотопам, средам обитания или экотопам в соответствие с их морфологическими идиоадаптациями, особенностями поведения, биологии и экологии.
МАТЕРИАЛ И МЕТОДЫ
На основе результатов многолетних исследований морфологии, экологии, этологии и биологии 141 вида амфипод Черного и Азовского морей (Булычева, 1957; Мордухай-Болтовской и др., 1969; Василенко, 1974; Грезе, 1977, 1985; Маккавеева, 1979; Киселева, 1981; Petrescu, 1998; Sezgin et al, 2001; Гринцов, 2009, 2010, 2011а, 2011б, 2016, 2017, 2021, 2022а, 2022б, 2022 в; Grintsov, 2009, 2010, 2014, 2018, 2021а, 2021б; Grintsov, Sezgin, 2011; Uzunova, 2012; Kudrenko, 2016) выделялись экологические группы, экоморфы и жизненные формы амфипод этих водоемов.
Для обозначения данных категорий были использованы морфологические идиоадаптации, выработанные амфиподами в ответ на наиболее критические условия той среды (или сред) обитания, которую (или которые) эти амфиподы предпочитают.
Фотографии габитусов и деталей тела взрослых самцов и самок сделаны с использованием микроскопа Hitachi SU 3500. Контурные зоологические рисунки ряда видов взяты из литературных источников.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Рассмотрим особенности выделенных категорий. Весь последующий анализ идиоадаптации и на их основе обозначенных категорий, рассматривается по сравнению с гаммаридной жизненной формой (сем. Gammaridae: рода Gammarus, Echinogammarus, Chaetogammarus и др.), предположительно более древней, поскольку известны окаменелые останки амфипод, близких по морфологии к этому семейству (Arfianti et аl., 2018). Кроме того, представители данной жизненной формы менее специализированы, что условно указывает на это семейство как на предковую группу (Гурьянова, 1951). В качестве анализируемых морфологических элементов рассматриваются не любые морфологические элементы, а идиоадаптации, способствующие выживанию на фоне тех экстремальных условий среды обитания (или сред обитания), к которым эти экоморфы и жизненные формы наиболее приспособлены.
Рассмотрим выделенные категории. Анализировали только идиоадаптации в связи с определенными условиями среды обитания. Название элементов морфологии здесь и далее даны в соответствие с принятыми для этой группы (Гурьянова, 1951; Barnard, Karaman, 1991). Группа трубкожилов характеризуется способностью строить трубки из «паутинного шелка» (Гурьянова, 1951). От других амфипод их отличает ряд следующих признаков: коксальные пластинки слабо развиты (функция защиты брюшной части тела ослаблена, боковые щиты не выражены), оостегиты самок (выводковые пластинки) узкие и длинные, не образующие сплошной камеры (Гурьянова, 1951). К этим признакам можно добавить несколько особенностей. Тело часто более округлое в поперечном сечении или сплющенное в спино-брюшном направлении. Кутикула в спинной части сегментов тела без зубцов или шипов (отсутствует дополнительная функция защиты тела от хищников); переоподы вооружены «паутинными» железами и поэтому определенные членики утолщены.
- трубкожилов, зарегистрированных в Черном и Азовском морях, можно обозначить три жизненных формы (рис. 1).
Тело амфипод монокорофиидной жизненной формы (рис. 1 А) уплощено сверху вниз; четвертые членики стебельков 2-й пары антенн у самцов вооружены зубцами (имеет место
39
Гринцов В. А.
![]()

Рис. 1. Внешний вид представителей монокорофиидной (А) ампитоидной (Б) и ампелисцидной жизненных форм трубкожилов
- – Monocorophium insidiosum (Crawford, 1937); Б – Ampithoe ramondi Leach, 1814; В – Ampelisca diadema (Costa, 1853). Масштабная литейка 1 мм.
добавление функции по (Северцов, 1949), а именно функции захвата, рис. 1 А, «А 2, зубцы»). Гнатоподы обеих пар с рядами длинных перистых щетинок, расположенных в виде ловчей сети (рис. 2 А, пища собирается по типу фильтрации).
Коксальные пластинки не смыкаются, слабо развиты (рис. 1 А, функция защиты брюшка
- боков ослаблена). Плеоподы короткие (рис. 2 Б, указаны стрелкой) не вытянуты как у других групп амфипод, что возможно отражается на эффективности плавания. Уроподы 3-й пары короткие и широкие, с почти округлыми ветвями (рис. 2 В, указаны стрелкой), без крючков или развитых шипов, что указывает на отсутствие использования их как зацепа за какие-либо предметы. Особи монокорофиидной формы избегают прибойных участков, предпочитая места с накоплением детрита – бухты, лагуны, эстуарии рек. В Черном и Азовском морях в эту форму входят следующие рода: Chelicorophium (7 видов), Corophium (2), Monocorophium
(2), Medicorophium (1), Crassicorophium (1).
Тело амфипод ампитоидной жизненной формы (рис. 1 Б) сжато с боков или близко к округлому в поперечном сечении. Коксальные пластинки 1–4 хорошо развиты и образуют щит, закрывающий брюшную часть с боков. Антенны 1-й и 2-й пар выполняют у одних видов сенсорную функцию, у других к ней добавляется функция собирания частиц с помощью плотных групп щетинок. Когти переопод морфологически однотипны, в виде крючков, часто дополнительно дистальный конец переона усилен шипами или зубцами рядом с креплением когтя (рис. 2 Г, указаны стрелками, функция зацепа за субстрат усилена). Иногда и когти усилены дополнительными шипами, например, у Ericthonius difformis (Spence Bate, 1857). Базиподиты 6-й и 7-й пар по форме однотипны. Уроподы 3-й пары у одних видов по типу гаммаридной формы, но ветви менее вооружены и менее развиты (рис. 2 Д, указаны стрелками). У других морфологически другие: стебельки короткие, ветви с крючками или мощными шипами (рис. 2 Ж, крючок указан стрелкой), что функционально выражается в смене роли рулей при плавании (роль рулей при плавании по (Barnard, Karaman, 1991), характерных для гаммаридной формы (рис. 10 К), на роль зацепа за элементы окружающей среды. Следует уточнить, что ветви 3-й пары уропод по типу рулей вытянутые, уплощенные или округлые в сечении, снабженные щетинками и шипами (рис. 10 К). Ветви уропод 3-й пары по типу зацепа короткие, конические, вооруженные развитыми зубцами, шипами и/или щетинками преимущественно терминально (рис. 2 Ж). Амфиподы ампитоидной жизненной формы предпочитают скопления водорослей в перифитоне и на дне. В Черном и Азовском морях в эту форму входят рода Ampithoe (1 вид), Cheirocratus (1), Cymadusa (1), Ericthonius
(3), Jassa (3), Pleonexes (1), Plumulojassa (1), Megamphopus (1), Photis (1), Microdeutopus (5), Microprotopus (2), Centraloecetes (1), Leptocheirus (1).
-
- амфипод ампелисцидной жизненной формы (рис. 1 В) глаза рудиментарны, в виде отдельных маленьких линз (функция зрения ослаблена). Коксальные пластинки 1–4 хорошо развиты (функция защиты брюшка усилена с боков, выражены боковые щиты). Переоподы 4-й пары с длинным тонким когтем (рис. 2 З, коготь указан стрелкой) и рядами развитых
40
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
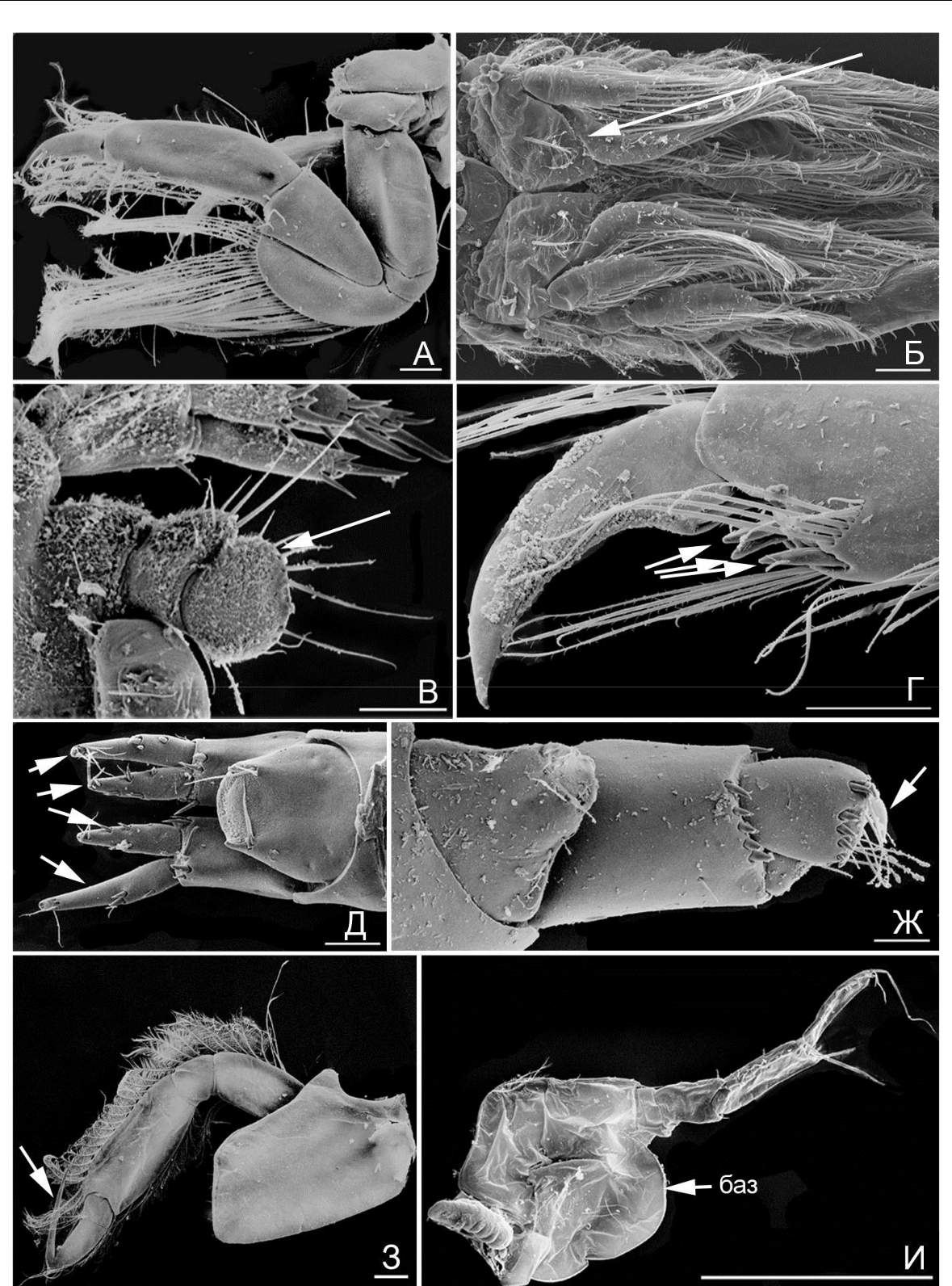
Рис. 2. Элементы морфологии амфипод
- – гнатопод 2, M. insidiosum; Б – плеоподы (стрелка), M. insidiosum; В – уросом и уропод 3 (стрелка), M. insidiosum; Г – A. ramondi, переопод 7, коготь и дистальная часть проподуса (стрелкой указаны шипы); Д – M. gryllotalpa, телсон и уроподы 3 (ветви указаны стрелкой); Ж – A. ramondi, тельсон и уропод 3; (стрелкой указан зубец); З – A. sevastopoliensis, переопод 4 (стрелкой указан коготь); И – A. diadema, переопод 6 («баз» – базиподит). Масштабная линейка 0,1 мм.
41
Гринцов В. А.
![]()
щетинок различного типа (простые, перистые, гребнистые), что указывает на использование этой пары конечностей для чистки тела и собирания частиц, возможно для собирания пищи. Базиподиты 6-й и 7-й пар переопод разной формы (рис. 2 И «баз»; рис. 3 А, «баз»). Когти переопод 6-й пары с группой дополнительных шипов (рис. 3 Б, указаны стрелками, функция захвата усилена, возможно добавление функции распределения «шелка» при постройке трубок).
Когти переопод 7-й пары каплевидной формы (рис. 3 А «ког», смена функции захвата на функцию упора). Уроподы 3-й пары с вытянутыми ветвями (рис. 3 В, ветви указаны стрелками), по типу рулей, но шипы и щетинки более редкие, чем у представителей гаммаридной жизненной формы. Особи ампелисцидной формы обитают почти исключительно среди детрита на рыхлом грунте. В Черном и Азовском морях в эту форму входит род Ampelisca (4 вида).
-
- амфипод-перфораторов выделена одна бианколинидная жизненная форма, внешний вид которой представлен на рис. 4.
Тело амфипод бианколинидной жизненной формы в поперечном сечении близко к округлому (рис. 4 А, Б); голова полушарообразная (рис. 4 А). Коксальные пластинки слабо развиты (рис. 4 А, Б, функция защиты брюшка ослаблена, за счет уменьшения коксальных пластинок, боковые щиты не выражены, функцию защиты при этом берут на себя ходы внутри перфорируемого объекта). Когти и проподальные членики вблизи когтей у некоторых видов (напр. B. algicola) могут быть усилены дополнительным вооружением (рис. 3 Г, шипы указаны стрелкой, функция зацепа усилена за счет усиления вооружения проподуса вблизи крепления когтя). Уроподы 3-й пары дополнительно вооружены (рис. 3 Д, шипы указаны стрелкой, функция зацепа усилена за счет добавления шипов или зубцов). Особи обитают либо внутри столонов крупных бурых макрофитов (B. algicola), либо в ходах внутри древесины (C. terebrans). В Черном и Азовском морях в эту форму входят два вида –
B. algicola, C. terebrans.
Комменсалы были разделены на 3 жизненные формы (рис. 5).
Тело амфипод коломастигидной жизненной формы в поперечном сечении почти округлое (рис. 5 А). Антенны обоих пар короткие, жгутики слабо развиты (рис. 5 А, сенсорная функция ослаблена за счет снижения числа члеников жгутиков, вооруженных чувствительными органами). Коксальные пластинки узкие, не закрывают полностью брюшную часть переона с боков (рис. 5 А, функция защиты брюшка с боков ослаблена, не выражены боковые щиты). Переоподы близкие по длине, их когти короткие (функция зацепа когтями за предметы ослаблена). Ветви 3-й пары уропод вытянуты, сужаются дистально, по типу рулей, но с более редкими шипами и щетинками, чем у представителей гаммаридной формы (напр. у Gammarus, Echinogammarus и пр.). В Черном море в эту жизненную форму входит один вид – C. pusilla.
Тело амфипод леукотоидной жизненной формы (рис. 5 Б) сжато с боков. Антенны 1-й и 2-й пар почти равны по длине, со слабо развитыми жгутиками (рис. 3 Б, сенсорная функция ослаблена за счет снижения числа члеников жгутиков, вооруженных чувствительными органами). Коксальные пластинки 1–4 хорошо развиты, образуют боковые щиты. Гнатоподы обоих пар (особенно 1-й) с характерным развитым выростом на карпальном членике (рис. 5 Б, функция захвата предметов элементами гнатопод усилена). Когти переопод небольшие и слабо загнутые (рис. 5 Б, функция захвата субстрата ослаблена). Уроподы с ветвями по типу рулей, сужающимися дистально, вооружение шипами и щетинками относительно гаммаридной формы значительно редуцировано (вероятно, ослаблена функция рулей при плавании). В Черном море (исключительно прибрежье Турции) в эту форму входит L. spinicarpa.
Тело амфипод кардиофилидной жизненной формы (рис. 5 В) сжато с боков. Антенны 1-
- пары длиннее антенн 2-й пары (рис. 5 В, сенсорная функция антенн 1-й пары усилена за счет увеличения числа члеников жгутиков с чувствительными органами). Гнатоподы 1-й и 2-й пар
42
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей

Рис. 3. Элементы морфологии амфипод
- – A. sevastopoliensis, переопод 7 («баз» – базиподит; «ког» – коготь); Б – A. sevastopoliensis, переопод 6, коготь (стрелкой указаны шипы); В – A. sevastopoliensis, уропод 3 (стрелкой указаны ветви); Г – B. algicola, переопод 5, коготь и дистальная часть проподуса (стрелкой указан шип); Д – B. algicola, уропод 3 и тельсон (стрелкой указаны шипы на ветвях уроподов 3); Ж – C. baeri, гнатопод 1; З – C. baeri, гнатопод 2; C. baeri, уросом, уроподы (стрелками указаны ветви уропод 3). Масштабная линейка: А, В,
Д-И – 0,1 мм; Б, Г – 0,01 мм.
43
Гринцов В. А.
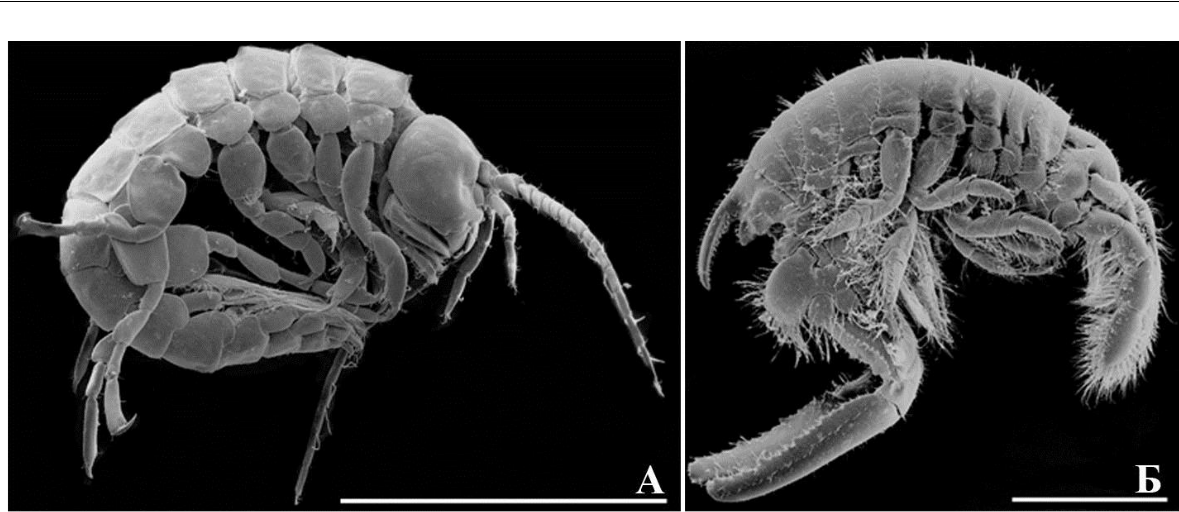
Рис. 4. Внешний вид особей бианколинидной жизненной формы
- – самка Biancolina algicola Della Valle, 1893; Б – самец Chelura terebrans Philippi, 1839. Масштабная линейка 1 мм.
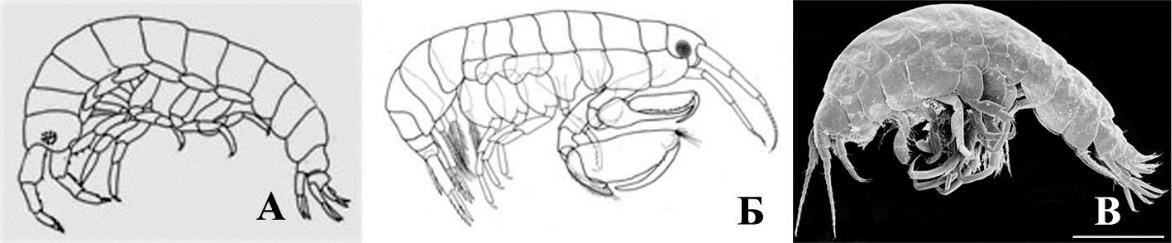
Рис. 5. Внешний вид представителей жизненных форм комменсалов: коломастигидной (А), леукотоидной (Б), кардиофилидной (В)
- – Colomastix pusilla Grube, 1861 (рис. по Barnard & Karaman, 1991); Б – Leucothoe spinicarpa (Abildgaard, 1789), (рис. по Krapp-Schickel, 1989); Г – Cardiophilus baeri G. O. Sars, 1896. Масштабная линейка 1 мм.
слабо развиты, разной формы (рис. 3 Ж, З, функция захвата предметов ослаблена). Коксальные пластинки 1–4 хорошо развиты (защита брюшной части переона с боков усилена). Уроподы 3-й пары с вытянутыми ветвями, но значительно меньше относительно тела и слабее вооружены, чем у гаммаридной жизненной формы (рис. 3 И, ветви указаны стрелками). Форма отмечена как комменсал двустворчатых моллюсков семейства Cardiidae, но также регистрируется в сообществах бентоса вне этих моллюсков. В Черном и Азовском морях эта форма представлена двумя видами рода Cardiophilus.
Мезопсаммофилы – амфиподы, зарывающиеся в песок (от греческого «meso» – внутри, «psammo» – песок, «phileo» – любить) были разделены на две жизненные формы (рис. 6).
Тело амфипод батипореидной жизненной формы (рис. 6 А) сужается от абдомена к голове (функция проникновения в песок облегчена). Между 1-м и 2-м члениками стебелька 1-
- пары антенн имеется перегиб на 90°, а сам дистальный участок 1-го членика с усиленным выростом (рис. 7 А, защита жгутика антенны усилена, указано стрелкой).
Коксальные пластинки 1–5 хорошо развиты, перекрываются, с венчиками длинных щетинок по краям (рис. 6 А, функция защиты брюшка с боков усилена, выражены боковые щиты). Гнатоподы 2-й пары без когтя, обильно снабжены группами длинных щетинок, что указывает на их собирающую функцию, но не на захват пищи обычным способом с помощью когтя (рис. 7 Б). Членики переопод значительно расширены, роющего типа (рис. 7 В, функция зарывания в песок усилена), вооружены множеством шипов и щетинок; когти переопод массивные, не загнутые (рис. 7 Г, указаны стрелкой, смена функции захвата на таковую
44
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
зарывания). Уроподы 3-й пары по типу рулей, с одной, но сильно развитой ветвью (рис. 7 Д, указана стрелкой), другая ветвь редуцирована до небольшой пластинки, менее вооружена щетинками и шипами по бокам, чем у представителей гаммаридной формы. Обитает ниже прибойной зоны, преимущественно глубже 10 м (Грезе, 1985). В Черном и Азовском морях отмечен один вид B. guilliamsoniana.
Амфиподы маеотикоидной жизненной формы (рис. 6 Б) с массивными антеннами, обильно вооруженными плотными группами щетинок (рис. 7 Ж), членики жгутиков антенн четковидные, массивные (функция проникновения в песок усилена, выражена функция защиты ротовых органов от песчинок). Коксальные пластинки 1–4 хорошо развиты, перекрываются, снабжены по свободным краям (вентрально, антеро-вентрально и постеро-вентрально) венчиками длинных щетинок (рис. 6, защитная функция брюшка с боков усилена, боковые щиты выражены). Гнатоподы и переоподы массивные, членики снабжены множеством длинных щетинок (рис. 7 З, функция зарывания в песок усилена), когти переопод развитые (рис. 7 З, функция захвата субстрата выражена). Уроподы 3-й пары по типу рулей, с развитыми ланцетовидными лопастями, обе пары лопастей (рис. 7 И, особенно наружная, указанная стрелкой) снабжены густой щеткой длинных перистых щетинок. В Черном и Азовском морях в маеотикоидную форму входит два рода и два вида понто-каспийских амфипод – Pontogammarus maeoticus (Sowinskyi, 1894) и Euxinia weidemanni (G. O. Sars, 1896).

Рис. 6. Внешний вид представителей батипореидной (А) и маеотикоидной (Б) жизненных форм мезопсаммофилов
- – Bathyporeia guilliamsoniana (Bate, 1857); Б – Pontogammarus maeoticus (Sowinskyi, 1894).
Масштабная линейка 1 мм.
Наконец, группа кривофилов (от греческого «kryvo» – прятаться, «phileо» – любить), предпочитает прятаться среди различных предметов (не используя указанные выше для других групп способы) – под камнями, среди водорослей, двустворчатых моллюсков, гидроидных полипов и пр. В данную группу входит большинство таксонов амфипод Черного
- Азовского морей. К этой группе относится капреллидная экоморфа, отличающаяся от всех остальных амфипод Черного и Азовского морей (семейство Caprellidae) и 9 жизненных форм. Амфиподы всех других экологических групп и оставшиеся амфиподы гаммаридной экологической группы по отношению к капреллидной экоморфе условно можно отнести к гаммаридной экоморфе. Гаммаридная экоморфа названа по семейству Gammaridae, входящему в нее, поскольку известны окаменелые остатки близкие к этому семейству (Arfianti et. al, 2018). Кроме того, Gammaridae менее специализированы по выбору субстрата относительно многих других семейств (Гурьянова, 1951), что условно указывает на это семейство как на предковую группу. В дальнейшем гаммаридная экоморфа анализироваться не будет, поскольку в нее входят все остальные амфиподы Черного и Азовского морей (кроме Caprellidae), которые будут рассматриваться подробнее по жизненным формам.
Капреллидная экоморфа (рис. 8 А, Б) отличается от гаммаридной экоморфы палочковидным телом, олигомеризацией сегментов тела, цефализацией, редукцией придатков – переопод, плеопод, уропод, а также усилением проподального членика и когтя.
45
Гринцов В. А.
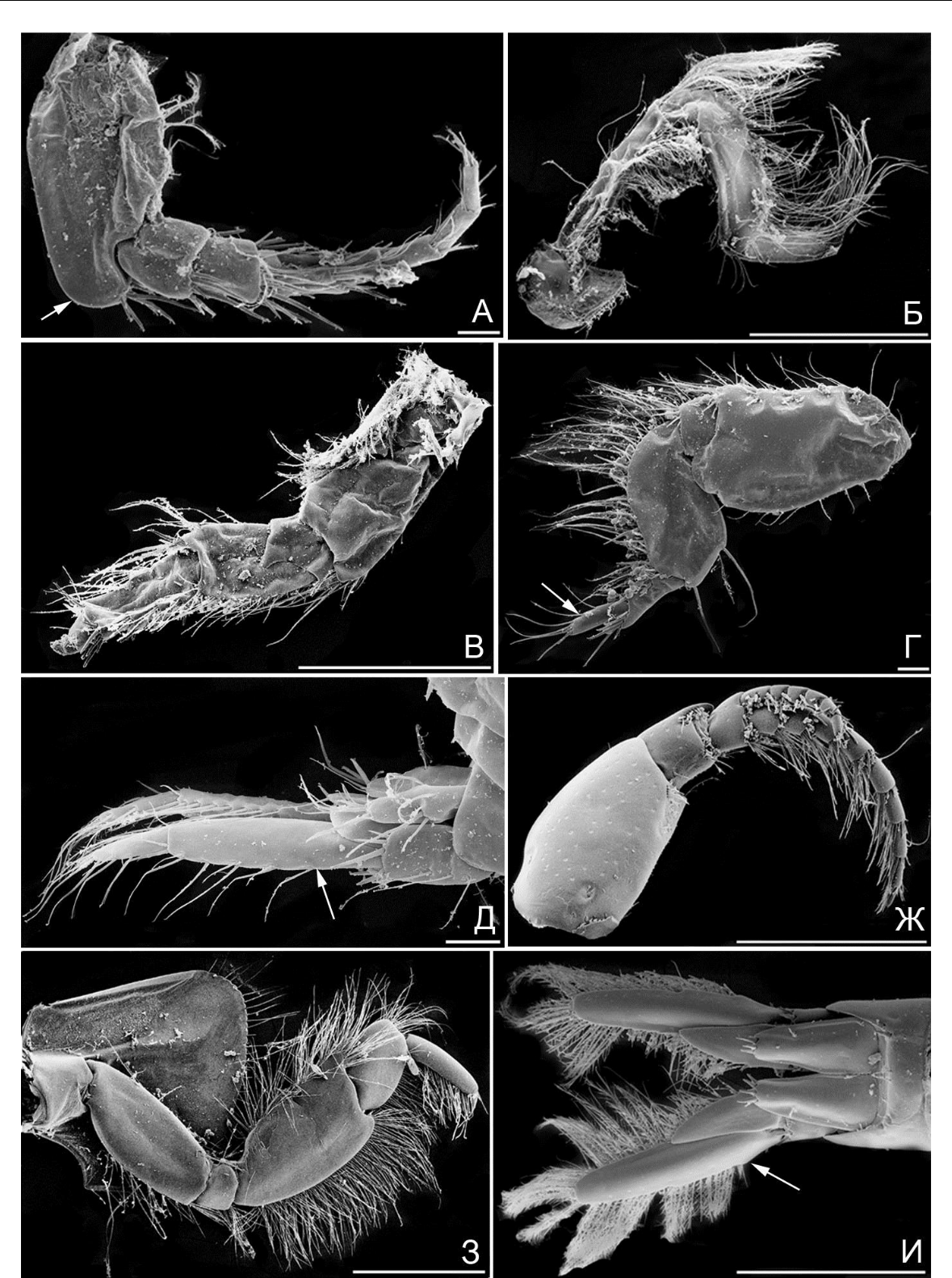
Рис. 7. Элементы морфологии амфипод
- – B. guiliamsoniana, антенна 1 (стрелкой указан выступ 1-го членика стебелька); Б – B. guiliamsoniana, гнатопод 2; В – B. guiliamsoniana, переопод 7; Г – B. guiliamsoniana, переопод 6 (стрелкой указан коготь); Д – B. guiliamsoniana, уроподы 3 (стрелкой указана наружная ветвь уропод 3); Ж – P. maeotica, антенна 1; З – P. maeotica, переопод 3; И – P. maeotica, уроподы 3 (стрелкой указана наружная ветвь). Масштабная линейка 1 мм.
46
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
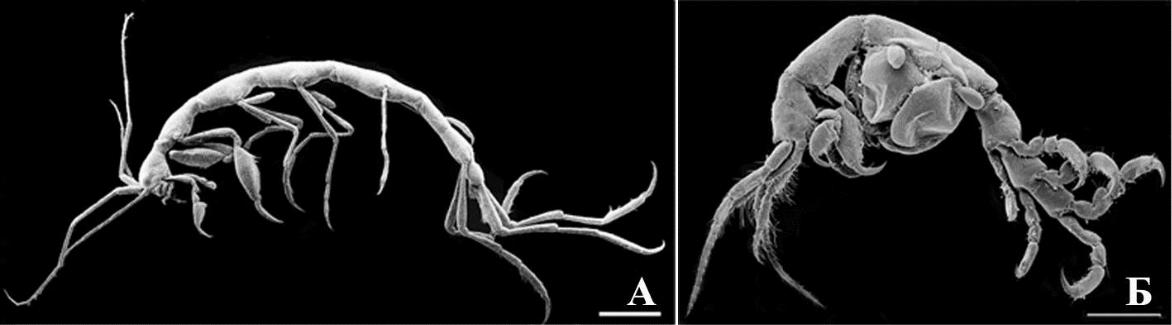
Рис. 8. Внешний вид представителей капреллидной экоморфы кривофилов
- – Phtisica marina Slabber, 1769; Б – Caprella liparotensis Haller, 1879. Масштабная линейка 1 мм.
Рассмотрим 9 жизненных форм кривофилов. Тело амфипод дексаминидной жизненной формы (рис. 9 А) с зубцами, либо валиками на спине (рис. 9 А, функция защиты от хищников усилена).

Рис. 9. Внешний вид представителей дексаминидной (А) хиалидной (Б) и гаммаридной жизненных форм кривофилов
- – Dexamine spinosa (Montagu, 1813); Б – Apohyale crassipes (Heller, 1866); В – Gammarus subtypicus Stock, 1866. Масштабная линейка 1 мм.
Капреллидная экоморфа обитает преимущественно на водорослях и гидроидных полипах. Несмотря на специализацию по среде обитания, эта экоморфа широко распространена из-за наличия водорослей почти во всех биотопах и сообществах перифитона
- бентоса. В Черном море к данной экоморфе относятся рода Caprella (6 видов), Phtisica (1), Pseudoprotella (1).
Антенны длинные или снабжены множеством чувствительных органов (рис. 10 А, Gammarellus), что указывает на усиление сенсорной функции. Гнатоподы однотипные по морфологии и сходные по размерам (рис. 10 Б, В). Коксальные пластинки 1–4 хорошо развиты, перекрываются (рис. 9 А, защита брюшка с боков усилена, боковые щиты выражены). Когти переопод развитые, загнутые (рис. 10 Г, указано стрелкой, функция захвата субстрата усилена). Уроподы 3-й пары с развитыми ланцетовидными ветвями (рис. 10 Д, ветви по типу рулей, указаны стрелками), однако вооружение менее обильно, по сравнению с представителями гаммаридной формы. Амфиподы дексаминидной формы характерны для водорослевых сообществ, преимущественно перифитона или различных скоплений водорослей, как в перифитоне, так и в бентосе. К дексаминидной жизненной форме в Черном
- Азовском морях относятся рода Apherusa (2 вида), Dexamine (3), Gammarellus (2), Tritaeta
(1), Nototropis (2).
Амфиподы эдицеротидной жизненной формы (рис. 11 А), с глазами, сдвинутыми к теменной части головы, или глаза отсутствуют (изменение или потеря функции зрения).
47
Гринцов В. А.
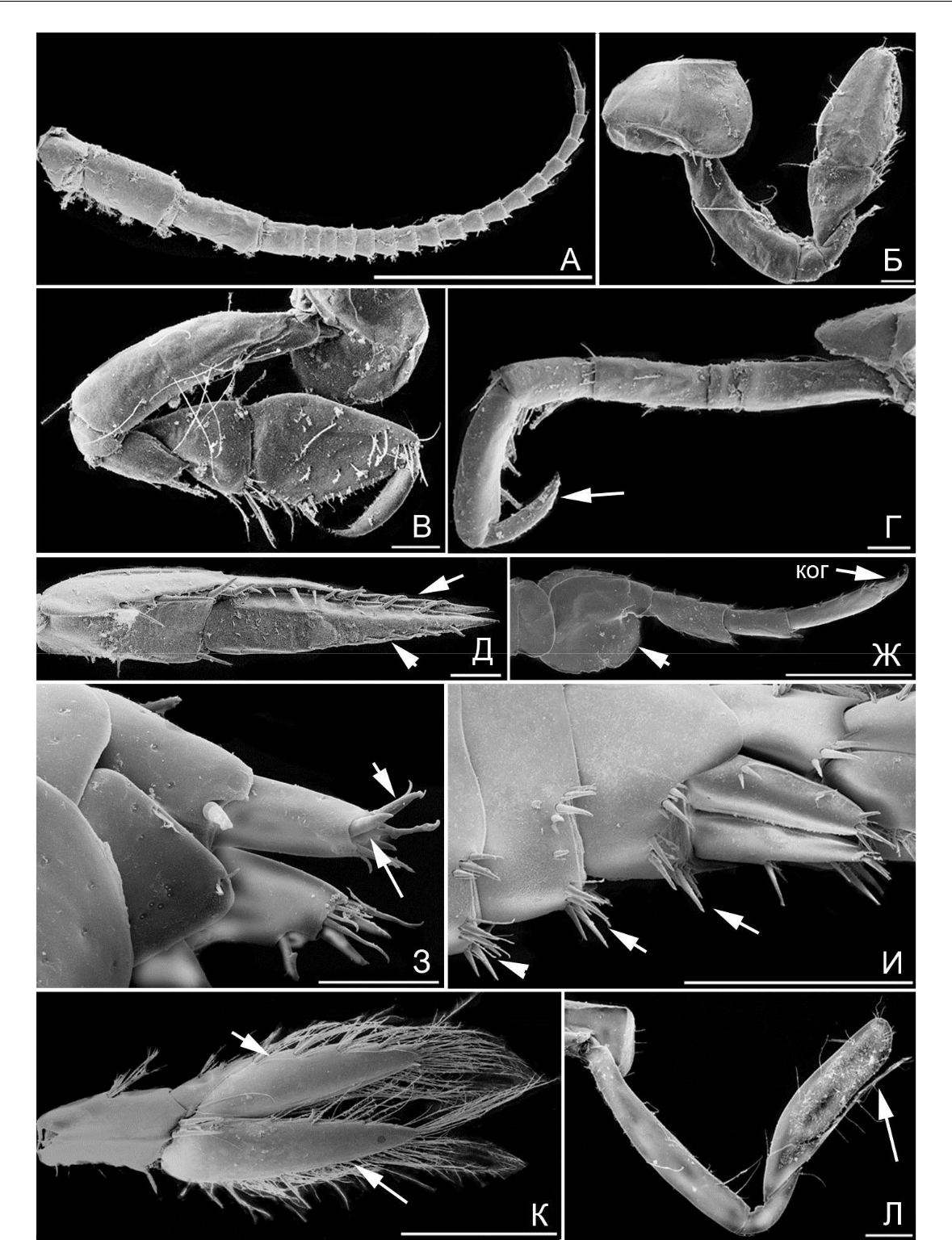
Рис. 10. Элементы морфологии амфипод
- – G. angulosus, антенна 1; Б – D. spinosa, гнатопод 1; В – D. spinosa, гнатопод 2; Г – D. spinosa,
переопод 4 (стрелкой указан коготь); Д – D. spinosa, уропод 3 (стрелками указаны выетви); Ж – A. perieri, переопод 7 (стрелкой указан базиподит, «ког» – коготь); З – A. perieri, тельсон и уропод 3 (стрелками указаны шипы ветви уропода 3); И – G. crinicornis, уросом вид латеро-дорзальный (стрелками указаны группы шипов и щетинок); К – G. aequicauda, уропод 3 (стрелками указаны ветви уропода 3); Л – P. longimanus, гнатопод 2 (стрелкой указан выступ карпуса). Масштабная линейка
0,1 мм.
48
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()

Рис. 11. Внешний вид представителей эдицеротидной (А) мегалуропиидной (Б) и талитридной жизненных форм кривофилов
- – Perioculodes longimanus (Spence Bate & Westwood, 1868); Б – Megaluropus massiliensis Ledoyer, 1976; В – Orchestia mediterranea Costa, 1853. Масштабная линейка 1 мм.
Коксальные пластинки 1–4 переона и эпимеральные пластинки 1-3 плеона образуют 2 боковых щита (рис. 11 А, функция защиты брюшка и плеопод усилена), частично отделенные зауженной частью. Карпальные членики гнатопод с выростами (рис. 10 Л, вырост указан стрелкой, функция захвата усилена). Переоподы 7-й пары чрезвычайно развиты относительно других пар, с прямым тонким и длинным когтем, не приспособленным для захвата субстрата, скорее он используется для упора на субстрат или погружения в рыхлый субстрат (рис. 11 А, смена функции захвата субстрата на функцию упора). Плеон удлиненный и массивный (рис. 11 А, функция плавания усилена), с развитыми плеоподами. Уроподы тонкие и длинные, дистально без шипов (рис. 12 А, часть ветвей указана стрелками, функция защиты уропод ослаблена за счет потери терминальных шипов).
Уроподы 3-й пары узкие, заостренные дистально (рис. 12 А, вероятно ослаблена функция рулей при плавании). Представители эдицеротидной формы обитают на рыхлых грунтах с примесью взвеси и детрита. Прямая форма когтей этой жизненной формы амфипод может затруднять их выживание в прибойной зоне, в том числе из-за ослабления или невозможности захвата за субстрат когтями во время штормов. В данную форму в Черном море входят рода Deflexilodes (2 вида), Perioculodes (1), Synchelidium (1).
Амфиподы хиалидной жизненной формы (рис. 9 Б) с хорошо развитыми коксальными пластинками 1–4 (рис. 6 Б, функция защиты брюшка с боков усилена, боковые щиты выражены) и расширенными базиподитами переопод с 5-й по 7-ю пары (рис. 10 Ж, базиподит указан стрелкой, функция защиты брюшка с боков усилена). Переоподы с развитыми загнутыми когтями и группами шипов в дистальной части проподального членика вблизи крепления когтя (рис. 10 Ж, «ког», функция захвата субстрата когтями усилена). Уроподы 3-
- пары маленькие, ветви короткие, субцилиндрические, с развитыми шипами на концах (рис. 10 З, шипы указаны стрелками, смена функции рулей на таковую упора при прыгании). Форма характерная для водорослевых сообществ, преимущественно перифитона. В Черном и частично Азовском морях к этой форме относятся рода Apohyale (3), Hyale (1), Micropythia
(1), Protohyale (2) Parhyale (2).
Амфиподы гаммаридной жизненной формы взяты за более древнюю, исходную Тело (рис. 9 В) вооружено в спинной части уросомы сочетанием шипов и щетинок или зубцов, шипов и щетинок (рис. 10 И, шипы частично указаны стрелками, выражена функция защиты от хищников). Коксальные пластинки 1–4 развитые, перекрывающиеся (рис. 9 В, функция защиты брюшка с боков усилена, боковые щиты выражены). Уроподы 3-й пары развиты, с ветвями в виде уплощенных вытянутых ланцетовидных лопастей (рис. 10 К, ветви указаны стрелками, ветви равны или одна существенно больше), снабженных различными щетинками и шипами по бокам (типичный тип рулей). Особи таксонов данной формы предпочитают скопления камней, водорослей и детрита на различных грунтах на дне. К этой форме относятся рода Amathillina (1), Chaetogammarus (3), Cheirocratus (1), Compactogammarus (1),
49
Гринцов В. А.

Рис. 12. Элементы морфологии амфипод
- – P. longimanus, уроподы (стрелками ветви); Б – M. massiliensis, коксальные пластинки 1-3; В – M. massiliensis, гнатопод 1, мерус-коготь; Г – M. massiliensis, тельсон, уроподы 3 (стрелками указаны ветви уропод); Д – O. gammarellus, антенна 1 (указана стрелкой); Ж – O. gammarellus, гнатопод 1, проподус и коготь; З – O. gammarellus, переопод 3, коготь (указан стрелкой); И – S. monoculoides, тельсон и уропод 3 (указан стрелкой). Масштабная линейка 0,1 мм.
50
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
Dikerogammarus (3), Echinogammarus (3), Euxinia (1), Gammarus (4), Gmelina (1), Gmelinopsis
(1), Iphigenella (3), Yogmelina (1), Kuzmelina (1), Melita (2), Niphargogammarus (1), Niphargoides
(1), Obessogammarus (2), Pandorites (1), Paraniphargoides (1), Pontogammarus (частично, 2 вида), Shablogammarus (1), Stenogammarus (5), Turcogammarus (1), Uroniphargoides (1).
Амфиподы мегалуропиидной жизненной формы (рис. 11 Б) с развитыми глазами, заходящими за боковые части головы (функция зрения усилена). Размеры коксальных пластинок специфически чередуются – вторая и четвертая больше, чем первая и третья (рис. 12 Б, усилена защита отдельных участков брюшной части переона). Гнатоподы обеих пар сходные морфологически, слабо развиты, без выраженного приспособления к захвату (рис. 12 В, не выражен край ладони проподуса со стороны когтя). Когти переопод вытянутые, прямые (функция захвата когтями субстрата ослаблена). Уроподы 3-й пары с развитыми листовидными широкими ветвями (рис. 12 Г, ветви указаны стрелками, по типу рулей). Представители этой формы обитают на рыхлых грунтах с примесью взвеси и детрита. Прямая форма когтей данной формы может затруднять выживание особей в прибойной зоне из-за ослабления захвата когтями за субстрат во время штормов. В мегалуропиидную форму в Черном море входят два вида из рода Megaluropus.
Амфиподы талитридной жизненной формы (рис. 11 В) с чрезвычайно маленькими и короткими антеннами 1-й пары (рис. 12 Д, антенна 1 указана стрелкой, сенсорная функция ослаблена за счет сокращения размеров и числа члеников жгутиков с чувствительными органами). Все придатки вооружены крепкими шипами, без щетинок. Все коксальные пластинки хорошо развиты (рис. 11 В, функция защиты брюшка с боков усилена). Гнатоподы 1-й пары самцов и самок маленькие, с рудиментарным краем со стороны когтя, сам коготь уменьшен в размерах (рис. 12 Ж, функция захвата предметов ослаблена). Когти переоподов мощные, загнутые (рис. 12 З, коготь указан стрелкой, функция захвата субстрата усилена). Уроподы 3-й пары маленькие, с уменьшенными короткими ветвями, близкие к конической форме, отсутствует щетка из щетинок разного типа, остаются только шипы (смена функции рулей при плавании на функцию элемента упора при прыгании). Данная форма исключительно наземная, особи обитают среди выбросов водорослей на пляжах. В Черном море в данную форму входят рода Britorchestia (1 вид), Cryptochestia (1), Orchestia (4), Platorchestia (1), Speziorchestia (1), Talitrus (1).
Амфиподы харпиниидной жизненной формы (рис. 13 А) без глаз (в Черном море), либо глаза разной формы у разных таксонов, относящихся к этой жизненной форме (преим. семейство Phoxocephalidae).

Рис. 13. Внешний вид представителей харпиниидной (А) стенотоидной (Б) и лизианассидной жизненных форм кривофилов
- – Harpinia crenulata (Boeck, 1871), по Karaman, 1993; Б – Stenothoe monoculoides (Montagu, 1815); В –
Orchomene humilis (Costa, 1853). Масштабная линейка 1 мм.
Над антеннами имеется характерный широкий козырек (функция защиты антенн усилена). Сами антенны короткие, сходные по длине (сенсорная функция ослаблена за счет сокращения числа члеников жгутиков с чувствительными органами). Переоподы 6-й пары значительно массивнее и длиннее остальных переопод; базиподиты переопод 6-й и 7-й пар разной формы. Когти переопод почти прямые (функция захвата субстрата ослаблена). Уроподы 3-й пары с ветвями, сужающимися дистально, ветви не ланцетовидные, по типу
51
Гринцов В. А.
![]()
рулей, вооружение щетинками частично редуцировано относительно гаммаридной формы. Представители данной жизненной формы обитают на илистых грунтах с примесью детрита. Прямая форма когтей харпиниидной формы может затруднять выживание особей в прибойной зоне из-за ослабления захвата за субстрат во время штормов. В Черном море харпиниидная жизненная форма представлена двумя видами рода Harpinia, обитающими а прибрежье Турции на глубине около 70 м.
Тело амфипод стенотоидной жизненной формы (рис. 13 Б), близко по контуру к овальному. Третья и особенно четвертая коксальные пластинки чрезвычайно развиты и образуют мощные боковые защитные щиты для брюшка и частично переопод (рис. 13 Б). Уроподы 3-й пары с одной палочковидной ветвью, слабо вооруженной шипами (рис. 12 И, ветвь указана стрелкой, по типу рулей). Данная форма характерна для водорослей. В Черном море отмечены представители рода Stenothoe (2 вида).
Амфиподы лизианассидной жизненной формы (рис. 13 В) со специфической формой 1-й пары антенн: стебельки вздутые, их членики бочёнковидные (рис. 14 А, членики стебелька указаны стрелкой).
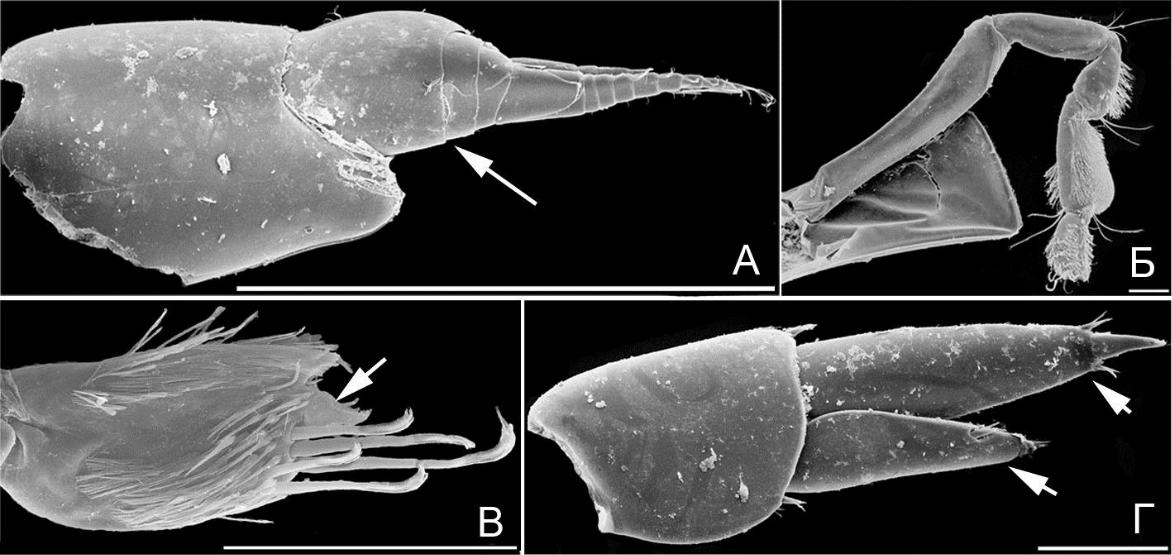
Рис. 14. Элементы морфологии амфипод
- – O. humilis, антенна 1 (стебелек указан стрелкой); Б – O. humilis, гнатопод 2; В – N. goesi reductus; гнатопод 2, проподус и коготь (коготь указан стрелкой); Г – O. humilis, уропод 3 (ветви указаны стрелкой). Масштабная линейка 0,1 мм.
Все коксальные пластинки хорошо развиты (рис. 13 В, функция бокового защитного щита усилена). Гнатоподы 2-й пары весьма специфические, тонкие; карпальный и проподальный членики близки по ширине и толщине, опушены специфически расположенной густой щеткой мелких, близких по длине щетинок (рис. 14 Б, преимущественно на задних поверхностях, вероятно выполняют роль чистящего органа), коготь маленький (рис. 14 В, коготь указан стрелкой, функция захвата предметов ослаблена). Уроподы 3-й пары с ветвями вытянуто-треугольными, без щетки щетинок (рис. 14 Г, ветви указаны стрелками, по типу рулей). У представителей данной формы нет выраженного предпочтения субстрата. В Черном море форма представлена родами Orchomene (1 вид) и Nannonyx (2).
- итоге, в результате исследований разработана общая схема выделенных экологических групп, экоморф и жизненных форм амфипод Черного и Азовского морей представленная в виде дендрограммы на основе экологии и морфологии на рисунках 15, 16.
52
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
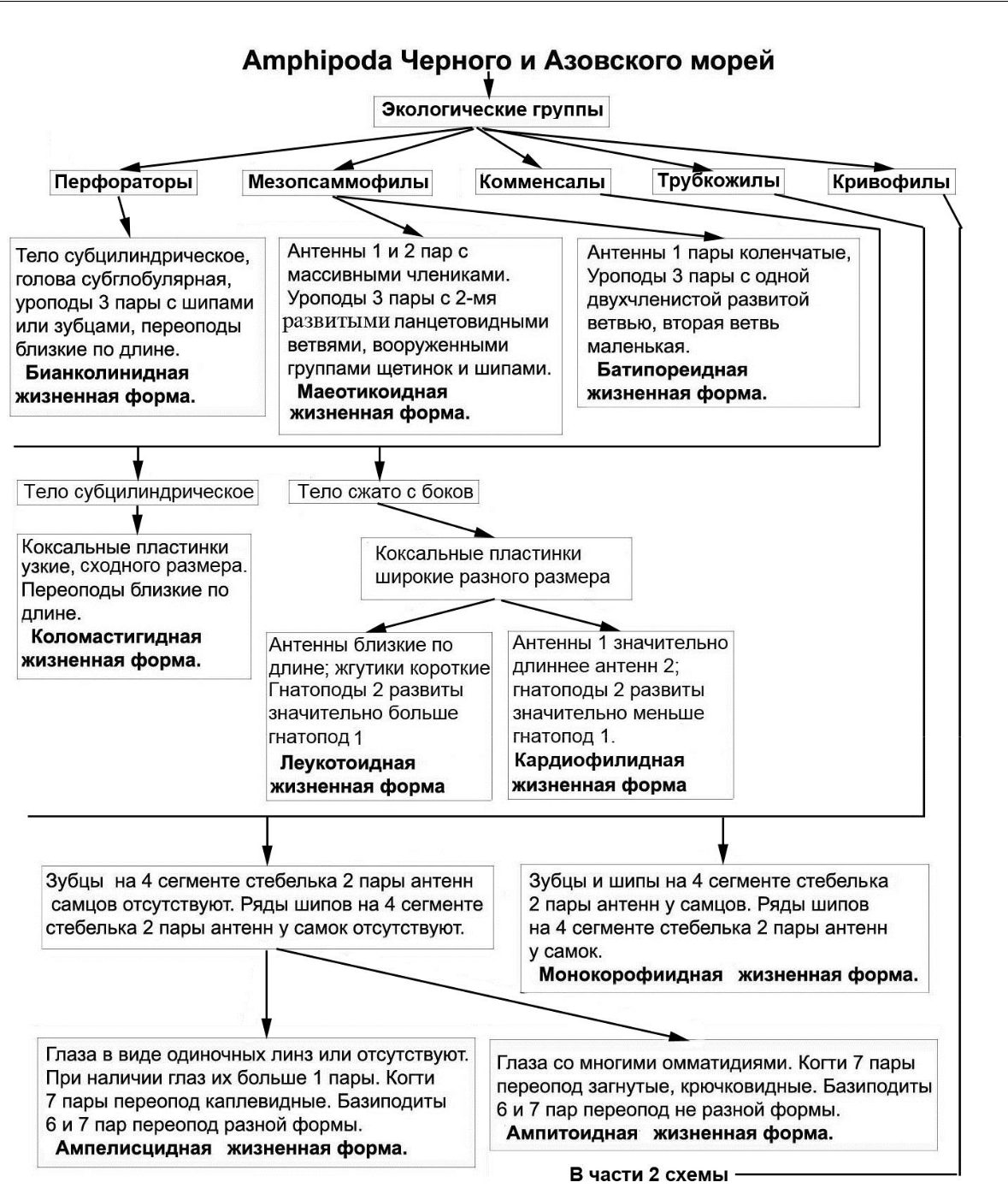
Рис. 15. Первая часть общей схемы выделения экологических групп, экоморф и жизненных форм амфипод Черного и Азовского морей
Данная схема позволяет проанализировать, какие вышеописанные идиоадаптации и другие морфологические преобразования способствовали сформированному в настоящее время эколого-морфологическому комплексу амфипод в Черном и Азовском морях. Как следует из схемы, наибольшими морфологическими преобразованиями подверглись амфиподы капреллидной экоморфы. Далее выделяется группа, способная строить трубки (трубкожилы). Выделение других групп связано со спецификой скрываться от хищников и опасностей (перфораторы, кривофилы, мезопсаммофилы, комменсалы). Образование внутри
53
Гринцов В. А.
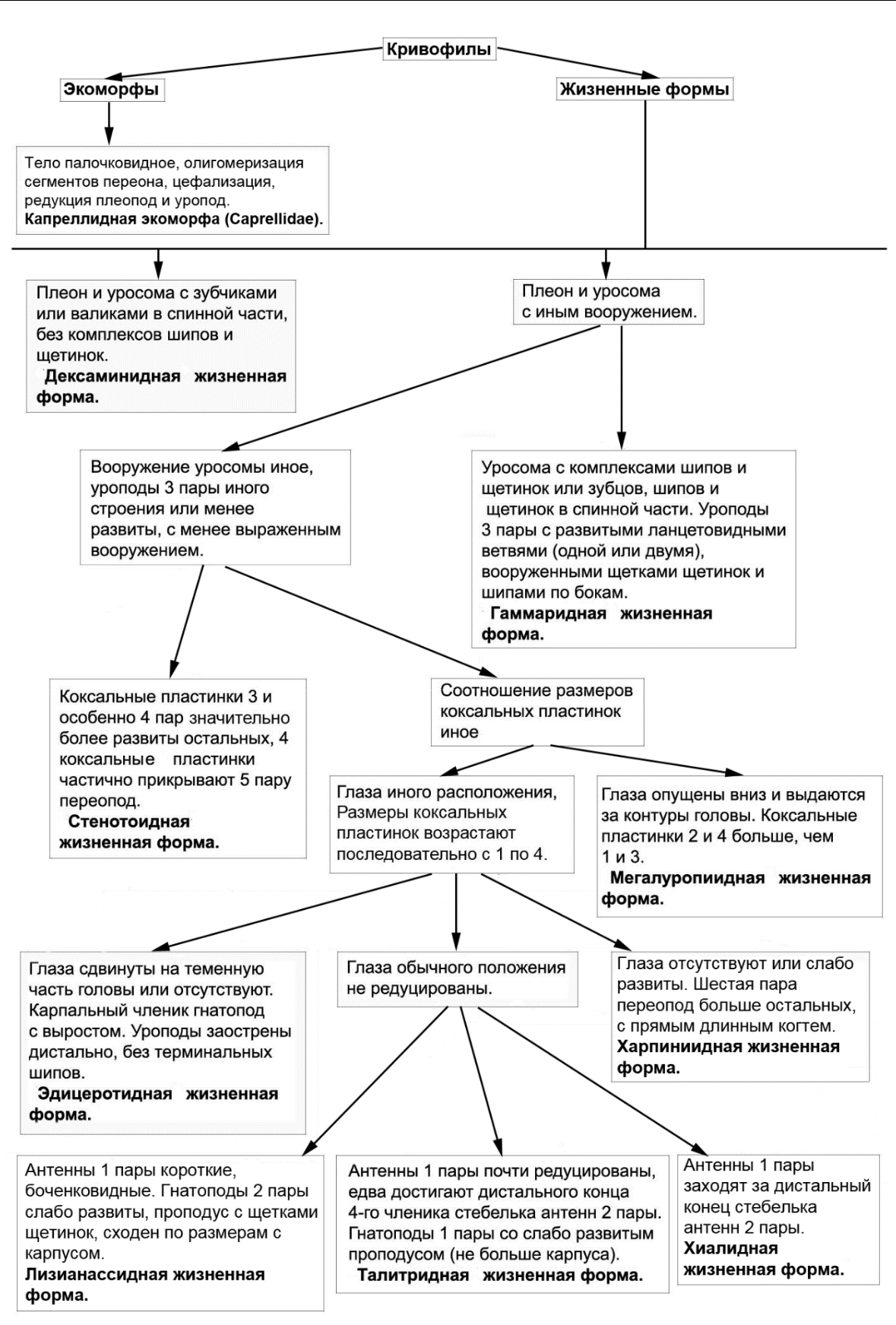
Рис. 16. Вторая часть общей схемы выделения экологических групп, экоморф и жизненных форм амфипод Черного и Азовского морей
54
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
экологических групп жизненных форм связано с появлением тех или иных идиоадаптаций, отчасти определяющих преимущественное выживание в тех или иных условиях.
Сообщества, биотопы, экотопы заселены различными экологическими группами, экоморфой и жизненными формами амфипод неравномерно. Наибольшее число жизненных форм и капреллидная экоморфа отмечены в перифитоне и водорослевых ассоциациях – монокорофиидная, ампитоидная, ампелисцидная жизненные формы трубкожилов; капреллидная экоморфа кривофилов; дексаминидная, хиалидная, гаммаридная, стенотоидная, лизианассидная жизненные формы кривофилов; бианколинидная жизненная форма перфораторов (B. algicola).
Выявленное разнообразие экологических групп и жизненных форм амфипод в данной экотопической группе (перифитон) отчасти объясняется сложной конфигурацией перифитона
- водорослевых ассоциаций, населенных множеством организмов. Так, в Черном море у берегов Крыма в перифитоне отмечено 275 видов макроорганизмов – макрофитов и беспозвоночных, относящихся к различным таксонам (Гринцов и др. 2004а, 2004б, 2005а, 2005б, 2005в; 2009). Подобное разнообразие способствует образованию множества микроусловий, обеспечивающих выживание разных амфипод и увеличивает пищевой спектр этих ракообразных. В результате к данной среде оказались приспособлены жизненные формы с разной адаптивной морфологией. Всего в перифитоне отмечено 42 вида этих ракообразных.
Следующая среда обитания относительно разнообразия экологических групп и жизненных форм – детрит и различные органические остатки на дне. В этой среде отмечены капреллидная экоморфа кривофилов; ампелисцидная жизненная форма трубкожилов;
гаммаридная, эдицеротидная, мегалуропиидная, харпиниидная, стенотоидная, лизианассидная жизненные формы кривофилов (Грезе, 1977, 1985; Киселева, 1980). Разнообразие жизненных форм и наличие капреллидной экоморфы в этой среде обитания объясняется отчасти богатой пищевой базой, включающей фрагменты растительного и частично животного происхождения. Кроме того, при достаточном слое детрита, мелкие особи различных жизненных форм могут спрятаться в нем, что способствует выживанию и как следствию увеличения разнообразия отмеченных категорий амфипод.
Несколько жизненных форм, представляющих две экологические группы, оказались связаны с различными беспозвоночными как комменсалы. Это коломастигидная, леукотоидная и кардиофилидная жизненные формы комменсалов, а также дексаминидная жизненная форма кривофилов – вид Tritaeta gibbosa (Spence Bate, 1862). В первую очередь в Черном море они связаны с губками, частично с двустворчатыми моллюсками семейства Cardiidae. Наличие нескольких жизненных форм сумевших приспособиться к комменсализму на вышеуказанных беспозвоночных, свидетельствует о возможности выбора разных путей адаптации к подобной среде.
Для остальных биотопов отмечено меньше экологических групп и жизненных форм, что свидетельствует об относительной ограниченности условий в плане выживания. Внутри древесины в ходах успешно выживает только 1 вид, относящийся к бианколинидной форме перфораторов (C. terebrans). В толще песка ниже прибойной зоны отмечен один вид мезопсаммофилов (B. guilliamsoniana). В толще песка зоны заплеска зарегистрированы 2 вида мезопсаммофилов (P. maeoticus, E. weidemanni). Толща песка в зоне супралиторали успешно заселена единственным видом талитридной формы Deshayesorchestia deshayesii (Audouin, 1826). В зоне заплеска галечно-песчанных пляжей отмечены некоторые представители гаммаридной формы кривофилов – Echinogammarus foxi Chellenberg, 1828; Echinogammarus ischnus (Stebbing, 1899); Chaetogammarus olivii (H.Milne Edwards, 1830); Echinogammarus karadagiensis Grintsov, 2009. Эти же виды, некоторые представители хиалидной жизненной формы кривофилов: Apohyale crassipes (Heller, 1866); Apohyale perieri (Lucas, 1849); Parhyale taurica Grintsov, 2009, а также один представитель ампитоидной жизненной формы – Plumulojassa ocia (Spence Bate, 1862), отмечены в зоне заплеска каменисто-валунных пляжей.
-
- сообществах бентоса по данным Киселевой (Киселева, 1980) отмечены представители ампелисцидной, ампитоидной, монокорофиидной жизненных форм трубкожилов, капреллидной экоморфы кривофилов, гаммаридной и стенотоидной жизненных форм
55
Гринцов В. А.
![]()
кривофилов. Всего в сообществах бентоса по данным (Киселева, 1980) отмечено 11 видов. Следует отметить, что почти все экологические группы, экоморфы и жизненные формы широко распространены в Мировом океане. Исключение составляют виды понто-каспийской реликтовой фауны, представляющие маеотикоидную жизненную форму маезопсаммофилов
- кардиофилидную жизненную форму комменсалов, ограниченные бассейнами Черного, Азовского и отчасти Каспийского морей. Общий список отмеченных видов в Черном и Азовском морях (с учетом литературных данных) представлен в таблице 1. Экологические группы, экоморфы и жизненные формы указаны только для видов.
Таблица 1
Список таксонов амфипод Черного и Азовского морей с указанием экологических групп,
экоморфы и жизненных форм
| Таксоны амфипод | Экологические | Экоморфы | Жизненные | |
| группы | формы | |||
| 1 | 2 | 3 | 4 |
| Ampeliscidae | |
| Ampelisca diadema (Costa, 1853) | трубкожилы |
| Ampelisca pseudosarsi Bellan-Santini | —//—//— |
| & Kaim-Malka, 1977 | |
| Ampelisca pseudospinimana Bellan- | —//—//— |
| Santini & Kaim-Malka, 1977 | |
| Ampelisca spinipes Boeck, 1861 | —//—//— |
| Ampithoidae | |
| Ampithoe ramondi Audouin, 1826 | —//—//— |
| Biancolina algicola Della Valle, 1893 | перфораторы |
| Cymadusa crassicornis (Costa, 1853) | трубкожилы |
| Pleonexes gammaroides Spence Bate, | —//—//— |
| 1856 | |
| Aoridae | |
| Microdeutopus algicola Della Valle, | —//—//— |
| 1893 | |
| Microdeutopus anomalus (Rathke, | —//—//— |
| 1843) | |
| Microdeutopus gryllotalpa Costa, | —//—//— |
| 1853 | |
| Microdeutopus stationis Della Valle, | —//—//— |
| 1893 | |
| Microdeutopus versiculatus (Spence | —//—//— |
| Bate, 1857) | |
| Atylidae | |
| Nototropis guttatus Costa, 1853 | кривофилы |
| Nototropis massiliensis (Bellan- | —//—//— |
| Santini, 1975) | |
| Bathyporeiidae | |
| Bathyporeia guilliamsoniana (Spence | мезопсаммофилы |
| Bate, 1857) | |
| Behningiellidae | |
| Cardiophilus baeri G. O. Sars, 1896 | комменсалы |
| Cardiophilus marisnigrae | —//—//— |
| Miloslawskaya, 1931 | |
| Calliopiidae | |
| Apherusa bispinosa (Spence Bate, | кривофилы |
| 1857) | |
| Apherusa chiereghinii Giordani- | —//—//— |
| Soika, 1949 |
гаммаридная
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
ампелисцидная
—//—//—
—//—//—
—//—//—
ампитоидная бианколинидная ампитоидная —//—//—
—//—//—
—//—//—
—//—//—
—//—//—
—//—//—
дексаминидная
—//—//—
батипореидная
кардиофилидная
—//—//—
дексаминидная
—//—//—
56
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
Таблица 1 (продолжение)
| 1 | 2 | 3 | 4 | |||
| Caprellidae | ||||||
| Caprella acanthifera Leach, 1814 | —// | —//— | капреллидная | —- | ||
| Caprella danilevskii Czerniavski, | —// | —//— | —// | —//— | —- | |
| 1868 | ||||||
| Caprella equilibra Say, 1818 | —// | —//— | —// | —//— | —- | |
| Caprella liparotensis Haller, 1879 | —// | —//— | —// | —//— | —- | |
| Caprella mitis Mayer, 1890 | —// | —//— | —// | —//— | —- | |
| Caprella rapax Mayer, 1890 | —// | —//— | —// | —//— | —- | |
| Phtisica marina Slabber, 1769 | —// | —//— | —// | —//— | —- | |
| Pseudoprotella phasma (Montagu, | —// | —//— | —// | —//— | —- | |
| 1804) | ||||||
| Cheirocratidae | ||||||
| Cheirocratus sundevallii (Rathke | трубкожилы | гаммаридная | ампитоидная | |||
| 1843) | ||||||
| Cheluridae | ||||||
| Chelura terebrans Philippi, 1839 | перфораторы | —// | —//— | бианколинидная | ||
| Colomastigidae | ||||||
| Colomastix pusilla Grube, 1861 | комменсалы | —// | —//— | коломастигидная | ||
| Corophiidae | ||||||
| Chelicorophium chelicorne (G. O. | трубкожилы | —// | —//— | монокорофиидная | ||
| Sars, 1895) | ||||||
| Chelicorophium curvispinum (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1895) | ||||||
| Chelicorophium maeoticum | —// | —//— | —// | —//— | —// | —//— |
| (Sowinsky, 1898) | ||||||
| Chelicorophium mucronatum (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1895) | ||||||
| Chelicorophium nobile (G. O. Sars, | —// | —//— | —// | —//— | —// | —//— |
| 1895) | ||||||
| Chelicorophium robustum (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1895) | ||||||
| Chelicorophium sowinskyi | —// | —//— | —// | —//— | —// | —//— |
| (Martynov, 1924) | ||||||
| Corophium orientale Schellenberg, | —// | —//— | —// | —//— | —// | —//— |
| 1928 | ||||||
| Corophium volutator (Pallas, 1766) | —// | —//— | —// | —//— | —// | —//— |
| Crassicorophium bonellii (H. Milne | —// | —//— | —// | —//— | —// | —//— |
| Edwards, 1830) | ||||||
| Crassicorophium crassicorne | —// | —//— | —// | —//— | —// | —//— |
| (Bruzelius, 1859); | ||||||
| Leptocheirus pilosus Zaddach, 1844 | —// | —//— | —// | —//— | ампитоидная | |
| Medicorophium runcicorne (Della | —// | —//— | —// | —//— | монокорофиидная | |
| Valle, 1893) | ||||||
| Monocorophium acherusicum (Costa, | —// | —//— | —// | —//— | —// | —//— |
| 1853) | ||||||
| Monocorophium insidiosum | —// | —//— | —// | —//— | —// | —//— |
| (Crawford, 1937) | ||||||
| Dexaminidae | ||||||
| Dexamine spiniventris (Costa, 1853) | кривофилы | —// | —//— | дексаминидная | ||
| D. spinosa (Montagu, 1813) | —// | —//— | —// | —//— | —// | —//— |
| D. thea Boeck, 1861 | —// | —//— | —// | —//— | —// | —//— |
| Tritaeta gibbosa (Spence Bate, 1862) | —// | —//— | —// | —//— | —// | —//— |
57
Гринцов В. А.
![]()
Таблица 1 (продолжение)
| 1 | 2 | 3 | 4 | |||
| Gammarellidae | ||||||
| Gammarellus angulosus (Rathke, | —// | —//— | —// | —//— | —// | —//— |
| 1843) | ||||||
| Gammarellus carinatus (Rathke, | —// | —//— | —// | —//— | —// | —//— |
| 1837) | ||||||
| Gammaridae | ||||||
| Amathillina cristata (G. O. Sars, | ||||||
| 1894) | —// | —//— | —// | —//— | гаммаридная | |
| Chaetogammarus placidus (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1896) | ||||||
| Chaetogammarus olivii (H. Milne | —// | —//— | —// | —//— | —// | —//— |
| Edwards, 1830) | ||||||
| Dikerogammarus villosus | ||||||
| (Sowinskyi, 1894) | —// | —//— | —// | —//— | —// | —//— |
| Dikerogammarus haemobaphes | —// | —//— | —// | —//— | —// | —//— |
| (Eichwald, 1841) | ||||||
| Dikerogammarus gruberi Mateus & | —// | —//— | —// | —//— | —// | —//— |
| Mateus, 1990 | ||||||
| Dikerogammarus istanbulensis | —// | —//— | —// | —//— | —// | —//— |
| Özbek, 2011 | ||||||
| Gmelina costata G. O. Sars, 1894 | —// | —//— | —// | —//— | —// | —//— |
| Gmelinopsis tuberculata G. O. Sars, | —// | —//— | —// | —//— | —// | —//— |
| 1896 | ||||||
| Yogmelina pusilla (G. O. Sars, 1896) | —// | —//— | —// | —//— | —// | —//— |
| Kuzmelina kusnezowi (Sowinskyi, | —// | —//— | —// | —//— | —// | —//— |
| 1894) | ||||||
| Shablogammarus subnudus (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 18961896) | ||||||
| Echinogammarus foxi (Schellenberg, | —// | —//— | —// | —//— | —// | —//— |
| 1928) | ||||||
| Echinogammarus ischnus (Stebbing, | —// | —//— | —// | —//— | —// | —//— |
| 1899) | ||||||
| Echinogammarus karadagiensis | —// | —//— | —// | —//— | —// | —//— |
| Grintsov, 2009 | ||||||
| Echinogammarus warpachowsky (G. | —// | —//— | —// | —//— | —// | —//— |
| O. Sars, 1894) | ||||||
| Gammarus aeqiucauda (Martynov, | —// | —//— | —// | —//— | —// | —//— |
| 1931) | ||||||
| Gammarus crinicornis Stock, 1966 | —// | —//— | —// | —//— | —// | —//— |
| Gammarus insensibilis Stock, 1966 | —// | —//— | —// | —//— | —// | —//— |
| Gammarus subtypicus Stock, 1966 | —// | —//— | —// | —//— | —// | —//— |
| Hyalidae | ||||||
| Apohyale crassipes (Heller, 1866) | —// | —//— | —// | —//— | хиалидная | |
| Apohyale perieri (Lucas, 1849) | —// | —//— | —// | —//— | —// | —//— |
| Apohyale prevostii (H. Milne | —// | —//— | —// | —//— | —// | —//— |
| Edwards, 1830) | ||||||
| Hyale pontica Rathke, 1836 | —// | —//— | —// | —//— | —// | —//— |
| Protohyale (Boreohyale) camptonyx | —// | —//— | —// | —//— | —// | —//— |
| (Heller, 1866) | ||||||
| Protohyale (Protohyale) schmidti | —// | —//— | —// | —//— | —// | —//— |
| (Heller, 1866) | ||||||
| Parhyale aquilina (Costa, 1857) | —// | —//— | —// | —//— | —// | —//— |
| Parhyale taurica Grintsov, 2009 | —// | —//— | —// | —//— | —// | —//— |
58
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
Таблица 1 (продолжение)
| 1 | 2 | 3 | 4 | |||
| Iphigenellidae | ||||||
| Iphigenella acanthopoda G. O. Sars, | —// | —//— | —//— | //— | гаммаридная | |
| 1896 | ||||||
| Iphigenella andrussowi G. O. Sars, | —// | —//— | —//— | //— | —// | —//— |
| 1894 | ||||||
| Iphigenella shablensis Carausu, 1943 | —// | —//— | —//— | //— | —// | —//— |
| Ischyroceridae | ||||||
| Centraloecetes dellavallei (Stebbing, | трубкожилы | —//— | //— | ампитоидная | ||
| 1899) | ||||||
| Ericthonius difformis H. Milne | —// | —//— | —//— | //— | —// | —//— |
| Edwards, 1830 | ||||||
| Ericthonius punctatus (Spence Bate, | —// | —//— | —//— | //— | —// | —//— |
| 1857) | ||||||
| Ericthonius rubricornis (Simpson, | —// | —//— | —//— | //— | —// | —//— |
| 1853) | ||||||
| Jassa marmorata Holmes, 1905 | —// | —//— | —//— | //— | —// | —//— |
| Jassa ocia (Spence Bate, 1862) | —// | —//— | —//— | //— | —// | —//— |
| Jassa pusilla (G.O. Sars, 1894) | —// | —//— | —//— | //— | —// | —//— |
| Kuriidae | ||||||
| Micropythia carinata (Spence Bate, | кривофилы | —//— | //— | хиалидная | ||
| 1862) | ||||||
| Leucothoidae | ||||||
| Leucothoe spinicarpa (Abildgaard, | комменсалы | —//— | //— | леукотоидная | ||
| 1789) | ||||||
| Lysianassidae | ||||||
| Nannonyx propinguus Chevreux 1911 | кривофилы | —//— | //— | лизианассидная | ||
| Nannonyx goesi reductus Greze, 1975 | —//— | //— | ||||
| Megaluropiidae | ||||||
| Megaluropus agilis Hoek, 1889 | кривофилы | —//— | //— | мегалуропиидная | ||
| Megaluropus massiliensis Ledoyer, | —// | —//— | —//— | //— | —// | —//— |
| 1976 | ||||||
| Melitidae | ||||||
| Melita palmata (Montagu, 1804) | —// | —//— | —//— | //— | гаммаридная | |
| Microprotopidae | ||||||
| Microprotopus longimanus Chevreux, | трубкожилы | —//— | //— | ампитоидная | ||
| 1887 | ||||||
| Microprotopus maculatus Norman, | —// | —//— | —//— | //— | —// | —//— |
| 1867 | ||||||
| Oedicerotidae | ||||||
| Deflexilodes gibbosus (Chevreux, | кривофилы | —//— | //— | эдицеротидная | ||
| 1888) | ||||||
| Deflexilodes griseus (Della Valle, | —// | —//— | —//— | //— | —// | —//— |
| 1893) | ||||||
| Perioculodes longimanus (Spence | —// | —//— | —//— | //— | —// | —//— |
| Bate& Westeood, 1868) | ||||||
| Synchelidium maculatum Stebbing, | —// | —//— | —//— | //— | —// | —//— |
| 1906 | ||||||
| Phoxocephalidae | ||||||
| Harpinia crenulata (Boeck, 1871) | —// | —//— | —//— | //— | харпиниидная | |
| H. dellavallei Chevreux, 1910 | —// | —//— | —//— | //— | —// | —//— |
| Photidae | ||||||
| Megamphopus cornutus Norman, | трубкожилы | —//— | //— | ампитоидная | ||
| 1869 | ||||||
| Photis longicaudata (Spence Bate & | —// | —//— | —//— | //— | —// | —//— |
| Westwood, 1862) | ||||||
59
Гринцов В. А.
![]()
Таблица 1 (продолжение)
| 1 | 2 | 3 | 4 | |||
| Pontogammaridae | ||||||
| Compactogammarus compactus (G. | кривофилы | —// | —//— | гаммаридная | ||
| O. Sars, 1895) | ||||||
| Euxinia sarsi (Sowinsky, 1898) | —// | —//— | —// | —//— | —// | —//— |
| Euxinia weidemanni (G. O. Sars, | мезопсаммофилы | —// | —//— | маеотикоидная | ||
| 1896) | ||||||
| Niphargogammarus intermedius | кривофилы | —// | —//— | гаммаридная | ||
| (Carausu, 1943) | ||||||
| Niphargoides corpulentus G. O. Sars, | —// | —//— | —// | —//— | —// | —//— |
| 1895 | ||||||
| Obesogammaruss crassus (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1894) | ||||||
| Obesogammaruss obesus (G. O. Sars, | —// | —//— | —// | —//— | —// | —//— |
| 1894) | ||||||
| Pandorites podoceroides G. O. Sars, | —// | —//— | —// | —//— | —// | —//— |
| 1895 | ||||||
| Paraniphargoides motasi (Carausu, | —// | —//— | —// | —//— | —// | —//— |
| 1943) | ||||||
| Pontogammarus abbreviatus (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1894) | ||||||
| Pontogammarus aestuarius | —// | —//— | —// | —//— | —// | —//— |
| (Derzhavin, 1924) | ||||||
| Pontogammarus maeoticus | мезопсаммофилы | —// | —//— | маеотикоидная | ||
| (Sovinskij, 1894) | ||||||
| Pontogammarus robustoides (G. O. | кривофилы | —// | —//— | гаммаридная | ||
| Sars, 1894) | ||||||
| Stenogammarus compressus (G. O. | —// | —//— | —// | —//— | —// | —//— |
| Sars, 1894) | ||||||
| Stenogammarus deminutus (Stebbing, | —// | —//— | —// | —//— | —// | —//— |
| 1906) | ||||||
| Stenogammarus kereuschi Derzhavin | —// | —//— | —// | —//— | —// | —//— |
| & Pjatakova, 1962 | ||||||
| Stenogammarus (Stenogammarus) | —// | —//— | —// | —//— | —// | —//— |
| macrurus (G. O. Sars, 1894) | ||||||
| Stenogammarus similis (G. O. Sars, | —// | —//— | —// | —//— | —// | —//— |
| 1894) | ||||||
| Turcogammarus aralensis (Uljanin, | —// | —//— | —// | —//— | —// | —//— |
| 1875) | ||||||
| Turcogammarus turcarum (Stock, | —// | —//— | —// | —//— | —// | —//— |
| 1974) | ||||||
| Uroniphargoides spinicaudatus | —// | —//— | —// | —//— | —// | —//— |
| (Carausu, 1943) | ||||||
| Stenothoidae | ||||||
| Stenothoe marina (Spence Bate, | —// | —//— | —// | —//— | стенотоидная | |
| 1856) | ||||||
| Stenothoe monoculoides (Montagu, | —// | —//— | —// | —//— | —// | —//— |
| 1813) | ||||||
| Talitridae | ||||||
| Britorchestia brito (Stebbing, 1891) | —// | —//— | —// | —//— | талитридная | |
| Cryptorchestia cavimana (Heller, | —// | —//— | —// | —//— | —// | —//— |
| 1865) | ||||||
| Deshayesorchestia deshayesii | —// | —//— | —// | —//— | —// | —//— |
| (Audouin, 1826) | ||||||
| Orchestia bottae H. Milne Edwards, | —// | —//— | —// | —//— | —// | —//— |
| 1840 | ||||||
60
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
Таблица 1 (продолжение)
| 1 | 2 | 3 | 4 | |||
| Orchestia gammarellus (Pallas, 1766) | —// | —//— | —// | —//— | —// | —//— |
| Orchestia mediterranea Costa, 1853 | —// | —//— | —// | —//— | —// | —//— |
| Orchestia montagui Audouin, 1826 | —// | —//— | —// | —//— | —// | —//— |
| Platorchestia platensis (Krøyer, | —// | —//— | —// | —//— | —// | —//— |
| 1845) | ||||||
| Speziorchestia stephenseni Cecchini, | —// | —//— | —// | —//— | —// | —//— |
| 1928 | ||||||
| Talitrus saltator (Montagu, 1808) | —// | —//— | —// | —//— | —// | —//— |
| Tryphosidae | ||||||
| Orchomene humilis (Costa, 1853) | —// | —//— | —// | —//— | лизианассидная | |
ВЫВОДЫ
-
- Черном и Азовском морях отмечено 5 экологических групп амфипод – трубкожилы, кривофилы, комменсалы, мезопсаммофилы, перфораторы. У трубкожилов выделены монокорофиидная, ампитоидная и ампелисцидная жизненные формы, идиоадаптации которых затронули следующие морфологические признаки: характер уплощения тела (с боков или сверху вниз), когти переопод, антенны 2-й пары, коксальные пластинки, переоподы, уроподы третьей пары. Сочетание данных идиоадаптаций специфично для каждой жизненной формы.
У кривофилов Черного и Азовского морей отмечена капреллидная экоморфа (семейство Caprellidae), отличающаяся палочковидным телом, олигомеризацией сегментов тела, цефализацией, редукцией придатков – переопод, плеопод, уропод, усилением проподальных члеников и когтей переопод.
Отмечено 9 жизненных форм кривофилов: дексаминидная, хиалидная, гаммаридная,
эдицеротидная, мегалуропиидная, талитридная, харпиниидная, стенотоидная, лизианассидная идиоадаптации которых затронули следующие морфологические элементы: рострум, глаза, вооружение спины, антенны 1 и 2 пар, коксальные пластинки, гнатоподы 1-й
- 2-й пар; уроподы, особенно 3-й пары, переоподы, когти переопод, вооружение уросомы. Сочетание морфологических идиоадаптаций специфично для каждой жизненной формы.
Бианколинидная жизненная форма перфораторов отличается цилиндрилизацией тела, полушарообразной головой, слабой развитостью коксальных пластинок, близкой длиной переопод. Идиоадаптации батипореидной и маеотикоидной жизненных форм мезопсаммофилов затронули антенны, гнатоподы, переоподы, уроподы 3-й пары. У коломастигидной, леукотоидной и кардиофилидной жизненных форм комменсалов идиоадаптации затронули антенны, коксальные пластинки, характер уплощенности тела, гнатоподы, переоподы, уроподы 3-й пары. Все вышеуказанные жизненные формы амфипод кроме кардиофилидной и маеотикоидной широко распространены в Мировом океане.
Наибольшее разнообразие экологических групп и жизненных форм в Черном море отмечено для перифитона и скоплений органических остатков (включая макрофиты) на дне. Меньше заселяется древесина, оказавшаяся в морской воде; заплеск галечно-песчанного пляжа; толща песка в зоне супралиторали пляжей.
Исследование выполнено в рамках темы госзадания «Исследование механизмов управления продукционными процессами в биотехнологических комплексах с целью разработки научных основ получения биологически активных веществ и технических продуктов морского генезиса» (№ гос. регистрации 121030300149-0)
61
Гринцов В. А.
![]()
Список литературы
Алеев Ю. Г. Жизненная форма как система адаптаций // Успехи современной биологии. – 1980. – Т. 90, вып.
- – С. 462–477.
Булычева А. И. Морские блохи морей СССР и сопредельных вод. – М-Л.: Изд-во АН СССР, 1957. – 185 с.
Василенко С. В. Капреллиды (морские козочки) морей СССР и сопредельных вод. – Л.: Наука, 1974. – 287 с.
Дедю И. И. Экологический энциклопедический словарь. – Кишинев.: Главная редакция Молдавской советской энциклопедии, 1990. – 406 с.
Димо Н. А., Келлер Б. А. В области полупустыни. – Саратов.: Изд. Саратовского губернского земства.
Почвенная. лаборатория, 1907. – 608 с.
Грезе И. И. Амфиподы Чёрного моря и их биология. – Киев.: Наук. думка, 1977. – 154 с.
Грезе И. И. Фауна Украины. Высшие ракообразные. Амфиподы. – Киев.: Наук. думка, 1985. – Т. 26, вып. 5. – 172 с.
Гринцов В. А., Мурина В. В., Евстегнеева И. К., Макаров М. А Сообщество обрастания на искусственном рифе в п. Курортное (Карадаг) // Карадаг. Гидробиологические исследования (сб. науч. тр. посвящ. 90-летию Карадаг науч. станции им. Т. И. Вяземского и 25-летию Карадаг. природ. Заповедника НАНУ). – Симферополь: 2004а. – Т. 2. – С. 152–165.
Гринцов В. А. Отряд амфиподы или разноногие раки (амфиподы) // Карадаг. Гидробиологические исследования (сб. науч. тр. посвящ. 90-летию Карадаг науч. станции им. Т. И. Вяземского и 25-летию Карадаг. природ. Заповедника НАНУ). – Симферополь, 2004б. – Т. 2. – С. 391–397.
Гринцов В. А., Костенко Н. С., Мурина В. В., Евстигнеева И.К. Биоразнообразие и структура сообщества акватории Карадагского природного заповедника // Заповедники Крыма: заповедное дело, биоразнообразие, экообразование: Матер. III науч. конф. (Симферополь, 22 апреля 2005.г.). – Симферополь, 2005а. – Часть 2. Зоология беспозвоночных, Зоология позвоночных. Экология. – С. 207–212.
Гринцов В. А., Мурина В. В., Евстигнеева И. К. Биоразнообразие и структура сообщества обрастания твердых субстратов Карадагского природного заповедника (Чёрное море) // Морской экологический журнал – 2005б. – Т. 4, № 3. – С. 37–47.
Гринцов В. А., Мурина В. В., Евстигнеева И. К. Новые сведения о биоразнообразии сообщества обрастаний твердых субстратов в прибрежной зоне Крыма // Науковi записки Тернопiльського нацiонального педагогiчного. унiверситету Серiя. Біологія. Спецiальний випуск: Гідроекологія – 2005в. – № 4 (27). – С. 54–55.
Гринцов В. А. Биоразнообразие и экология бокоплавов прибрежья Карадага // Карадаг – 2009 (сб. науч. Тр. посвящ. 95-летию Карадаг науч. станции и 30-летию Карадаг. природ. Заповедника НАНУ). – Севастополь, 2009б. – С. 361–365.
Гринцов В. А. Морфофункциональные параметры плавательных ножек плеопод (Амфипод) и некоторые моменты филогении и систематики // Бюллетень МОИП – 2010. – Т. 115, вып. 5. – С. 36–42.
Гринцов В. А. Морфологические различия видов рода Hyale (Amphipoda, Hyalidae) из Чёрного моря (прибрежье Крыма) // Весник Зоологии. – 2011а. – № 45 (5). – С. 447–455.
Гринцов В. А. Морфологические различия самок рода Orchestia и Platorchestia (Amphipoda, Talitridae) супралиторали Чёрного моря (Крым) // Зоологический журнал – 2011б. – Т. 90, № 2. – С. 143–148.
Гринцов В. А. Динамика структуры популяции двух видов бокоплавов рода Echinogammarus (Gammaridae, Amphipoda) из бухты Ласпи (Крым, Чёрное море) // Морской биологический журнал – 2016в. – Т. 1, № 3. – С. 22– 26.
Гринцов В. А. Биоразнообразие и динамика численности бокоплавов на коллекторах мидийной фермы (Севастополь, Черное море) // Морской биологический журнал – 2017. – Т 2, № 4. – С. 30–37.
Гринцов В. А. Амфиподы морских ферм прибрежья Севастополя (Черное море) // Водные биоресурсы и среда обитания – 2021. – Т. 4, № 4. – С. 27–35.
Гринцов В. А., Бондаренко Л. В., Тимофеев В. А. Новый вид амфиподы Melita Leach, 1814 (Crustacea, Amphipoda, Melitidae) для Азово-Черноморского бассейна // Российский журнал биологических инвазий – 2022а. –
- 5, № 1. – С. 41–54. https://doi.org/10.35885/1996-1499-15-1-41-54
Гринцов В. А. Амфиподы Черного моря: иллюстрированный атлас-определитель // ФИЦ «Институт биологии
южных морей им. А.О. Ковалевского РАН». – Севастополь: ФИЦ ИНБЮМ, 2022б. – 476 c.
Гурьянова Е. Ф. Амфиподы морей СССР. – Л.: Изд-во АН СССР, 1951. – 1029 с.
Киселева М. И. Бентос рыхлых грунтов Чёрного моря. – Киев, Наук. думка, 1981. – 168 с.
Маккавеева Е. Б. Беспозвоночные зарослей макрофитов Чёрного моря. – Киев, Наукова думка,. 1979. – 228 с. Мордухай-Болтовской Ф. Д., Грезе И. И., Василенко С. В. Отряд амфиподы или разноногие. Amphipoda // –
В кн.: Определитель фауны Чёрного и Азовского морей. – Киев, Наук. думка, 1969. – Т. 2. – С. 440–494.
Северцов А. Н. Морфологические закономерности эволюции. – М-Л.: Изд-во. АН СССР, 1949. – Т. 5. – С. 1–
536.
Формозов А. Н. Млекопитающие Северной Монголии по сборам экспедиции 1926 г. – М.: 1929. – 144 с. Arfianti T., Wilson S., Costello M. J. Progress in the discovery of amphipod crustaceans. – 2018. PeerJ 6:e5187; DOI
10.7717/peerj.5187
Barnard J. L., Karaman G. S. The families and genera of marine Gammaridean Amphipoda (except marine Gammaroids) // Records of the Australian Museum. – 1991. – Suppl. 13 (Parts 1–2). – P. 866.
62
Экологические группы, экоморфы и жизненные формы
амфипод (Crustacea, Amphipoda) Черного и Азовского морей
![]()
Grintsov V. A new amphipod species Echinogammarus karadagiensis sp. n. (Amphipoda, Gammaridae) from Crimean coasts (Black Sea, Ukraine) // Vestnik Zoologii. – 2009. – Vol. 43, N 2. – P. 23–26.
Grintsov V. On finding Dexamine thea (Amphipoda, Dexaminidae) in the Ukrainian territorial waters (Crimea, Black Sea) // Vestnik zoologii – 2010. – Vol. 44, N 3. – P. 281–283.
Grintsov V. Dynamyx of population structure of the Amphipod Dexamine thea (Dexaminidae, Amphipoda), a new species for the Black Sea in the coastal areas in Laspi Bay (Black Sea, Sevastopol) // Russian Journal of Biological Invasions. – 2014. – Vol. 5, N 1. – Р. 18–20.
Grintsov V. On finding of Monocorophium insidiosum Crawford, 1937 (Amphipoda, Corophiidae) in the coastal waters of Crimea (Black sea), a new species for this region // Marine biological journal. – 2018. – Vol. 3, N 2. – P. 33–39.
Grintsov V. А. First finding of Centraloecetes cf. neapolitanus (Schiecke, 1978) (Ischyroceridae, Amphipoda) in coastal zone of Sevastopol (Crimea, Black Sea) // Труды Карадагской научной станции им. Т. И. Вяземского – природного заповедника РАН. – 2021а. – № 2 (18). – С. 3–11.
Grintsov V. A. First finding of Caprella сf. equilibra Say, 1818 (Amphipoda, Capreliidae) in coastal zone south-west of Crimea (Black Sea) // Труды Карадагской научной станции им. Т. И. Вяземского – природного заповедника РАН. – 2021б. – № 4 (20). – С. 3–9.
Grintsov V., Sezgin M. Manual for identification of Amphipoda from The Black Sea // Sevastopol. Digit Print. – 2011. – 151 p.
Kudrenko S. A. Amphipod (Crustacea, Amphipoda) communities in the north-western part of the Black Sea // Vestnik zoologii – 2016. – Vol. 50 (5). – P. 387–394.
Petrescu I. Contribution to the knowledge of Amphipods (Crustacea: Amphipoda) from Romania. 7. Amphipods from Agigea (Black Sea) // Travaux du Museum d’Historie Naturelle “Grigore Antipa”. – 1998. – Vol. 15. – P. 51–73.
Sezgin M., Kocataş A., Katağan T., Amphipod Fauna of the Turkish Central Black Sea Region. // Turkish Journal of Zoology – 2001. – Vol. 25. – P. 57–61.
Uzunova S. Checklist of marine Amphipoda (Crustacea, Malacostraca) from the Bulgarian Black Sea area // Известия на съюза на учените – Варна, Серия «Морски науки». – 2012. – С. 72–79.
Grintsov V. A. Ecological groups, ecomorphs and life forms of amphipods (Crustacea, Amphipoda) of the Black Sea and the Sea of Azov // Ekosistemy. 2023. Iss. 33. P. 38–63.
On the basis of the analysis of morphology, biology, ethology, ecology of 141 species of amphipods of the Black Sea and the Sea of Azov, five ecological groups were identified: tubebuilders, perforators, commensals, kryvophiles (from the Greek «kryvo» – to hide, «phileo» – to love) and mesopsammophiles (from the Greek «meso» – inside, «psammo» – sand, «phileo» – to love). Moreover, ecomorphs and life forms are defined within these five ecological groups. The caprellid ecomorph (family Caprellidae) and nine life forms named after the taxa included in them are designated in the kryvophiles: hialid (genus Hyale), dexaminid (genus Dexamine), gammarid (genus Gammarus), edicerotid (family Oedicerotidae), megaluropiid (genus Megaluropus), talitrid (family Talitridae), harpiniid (genus Harpinia), stenotoid (genus Stenothoe), Lisianassid (family Lisianassidae). The life forms of all five ecological groups are not restricted to the taxa they are named after. Tubebuilders are divided into ampeliscid (genus Ampelisca), ampitoid (genus Ampithoe) and monocorophiid (genus Monocorophium) life forms. The perforators have one life form – biancolinid (genus Biancolina). The commensals have three life forms: cardiophilid (genus Cardiophilus), colomastigid (genus Colomastix) and leukotoid (genus Leucothoe). Two life forms were identified in mesopsammophiles – bathyporeid (genus Bathyporeia) and maeoticoid (species Pontogammarus maeoticus). This article analyzes ecological groups, ecomorphs and life forms, as well as their distribution in communities, biotopes, habitats or ecotopes in accordance with morphological idioadaptations of amphipods, ethology, ecology and biology.
Key words: Amphipoda, ecomorphs, ecological groups, live forms, Black Sea, Sea of Azov.
Поступила в редакцию 11.01.23
Принята к печати 05.02.23
63
