
SURGES OF NATURAL RECOVERY OF SCOTS PINE (PINUS SYLVESTRIS L.) AND CRIMEAN PINE (PINUS PALLASIANA D. DON): SYNCHRONICITY AND GENERAL PATTERNS
JOURNAL: «EKOSISTEMY», Issue 27, 2021
Publication text (PDF):Download
UDK: 582.475:574:630*231
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Saltykov A. N.
V.I. Vernadsky Crimean Federal University, Simferopol, Russia
TYPE: Article
DOI: https://doi.org/10.37279/2414-4738-2021-27-23-35
PAGES: from 23 to 35
STATUS: Published
LANGUAGE: Russian
KEYWORDS: Scots pine (Pinus sylvestris), Crimean pine (Pinus pallasiana), population, recovery surge, ecological niche, seedlings, seedage, undergrowth, young growth.
ABSTRACT (ENGLISH):
The study was carried out on the territory of the south-western segment of the Russian Plain in 2003–2020, so this research made it possible to identify a number of patterns typical for the process of natural recovery of pine forests (Pinus sylvestris L.). In particular, the obtained data indicate that population surges within the boundaries of the semiarid zone of the plain correlate with the climatic features of the region and are connected with the processes of Atlantic cyclonicity. The trail of the population surge, characteristic to steppe pine forests, is traced in the zone of coniferous-deciduous forests and on the southern borders of the plain, in the forest-steppe foothills of Crimea. The researchers confirmed the homogeneity of circulation processes and the course of the main climatic indicators. It is supposed to be one of the most probable reasons for the synchronicity of bursts of recovery on the vast territory of the south-western segment of the Russian Plain. Periodic repetitive population surges are aimed at restoration of the lost living space of pine forests. Autoregulation of the structure of undergrowth cenopopulations in accordance with the capacity of the renewal niche and a variety of ecological conditions allows each subsequent wave of renewal to master all potentially possible niches of the pine population space and is the basis for improving the structure and stability of the population.
ВВЕДЕНИЕ
На фоне общей тенденции потепления климата превышение среднемесячных температур в ряде регионов Российской Федерации влечёт за собой катастрофическое распространение лесных пожаров. В настоящее время сильнее всего от лесных пожаров страдают Дальневосточный и Сибирский федеральные округа. Для Русской равнины ближайшими во времени и хорошо известными примерами последствий атмосферных засух являются лесные пожары 1972 и 2010 годов (Вакуров, 1975; Мячкова, 1983, Переведенцев и др., 2011; Салтыков, 2019). Восстановление лесов искусственным путём в труднодоступных и малонаселённых регионах требует значительных и не всегда оправданных затрат. В тоже время лесной пожар можно рассматривать как условие сохранения популяционного пространства светлохвойных пород (Дмитриевский, 1928; Мелехов, 1944; Краснов, 1950; Тюрин, 1952; Ткаченко, 1955; Врадий, 1961; Вакуров, 1975; Одум, 1986; Синицын, 2008; Коба, 2017; Салтыков, 2019; Санников и др., 2019). Так, например, в начале столетия М. Е. Ткаченко, исследуя структуру лесного покрова Европейского Севера России установил, что в результате засушливых периодов на обширных пространствах Русской равнины сформировались большие массивы одновозрастных древостоев сосны. Пирогенное происхождение коренных сосняков, сложенных разобщёнными пространственными кластерами, по мнению учёного, не вызывает сомнений (Вакуров, 1975). Расширение списка «пожарных лет» для лесов Севера было продолжено академиком И. С. Мелеховым (1944). По мнению учёного, недостаточное увлажнение и следующая за ним засуха, являются причиной периодически повторяющихся масштабных по площади лесных пожаров (Мелехов, 1944; Ткаченко, 1955). Доминирование одновозрастных насаждений сосны следствие всплеска возобновления светлохвойных пород в границах пирогенного ряда. Исследованиями А. В. Тюрина установлено, что на обширных пространствах Русской равнины от Архангельской и Вятской губернии до Орловской, а ныне Брянской области, сосновые древостои появлялись синхронно во времени (Тюрин, 1952; Ткаченко, 1955). Аналогичные закономерности были выявлены учёными для южной покатости Русской равнины. Так, В. Д. Огиевский обращает внимание на согласованность процессов плодоношения и естественного возобновления сосняков на боровых террасах Днепра и Десны, в Полесье и лесостепной зоне равнины (Огиевский, 1966). Изучая естественное возобновление пристепных боров реки Северский Донец, П. П. Дмитриевский выявил волновой, пульсирующий характер процесса. Пики возобновления сосны, по сведениям учёного, пришлись на 1919 и 1926 годы. (Дмитриевский, 1928). Исследователь одним из первых установил, что всплеск возобновления можно одновременно наблюдать на горельниках и землях, выведенных из-под сельскохозяйственного пользования (Дмитриевский, 1928). Волна возобновления, по мнению П. П. Дмитриевского, коррелирует с большим количеством осадков, приходящих на боровую террасу Северского Донца в течение весенне-летнего сезона. Массовое появление самосева сосны в 1926 году в борах Чугуевской лесной дачи, расположенной на левом берегу Северского Донца, отражено в работах Г. Н. Высоцкого (1929). В том же 1926 году всплеск возобновления установлен исследователями Бузулукского бора (Краснов, 1950). Исследуя процессы естественного возобновления придонецких боров, Н. И. Врадий указывает на синхронность популяционных всплесков в бассейне Северского Донца и Днепра. Автор датирует всплеск возобновления в 1942–1943 годах, обращая внимание на то, что указанный период отличался избыточным увлажнением: «По метеорологическим данным за предшествующие 50 лет и сведениям из климатологической литературы установлены исключительно влажные годы, какими являлись: 1893, 1903, 1912, 1919, 1922, 1925, 1931–32 годах и анализируемый нами период 1941–1943 годов» (Врадий, 1975). Синхронное во времени появление подроста сосны на боровых террасах Донца, Днепра и Десны также было установлено С. С. Пятницким (1964). В рамках указанных исследований отчётливо прослеживается последовательность рассматриваемого процесса: засуха, лесной пожар, увеличение количества осадков, всплеск возобновления сосняков (Дмитриевский, 1928; Мелехов, 1944; Краснов, 1950; Врадий, 1961; Вакуров, 1975; Одум, 1986).
Вероятно, следует обратить внимание на существующую согласованность периодов естественного возобновления в границах семиаридной зоны Русской равнины с данными приведёнными А. Д. Вакуровым для северной её покатости. В частности, даты пожаров для Онежского лесхоза (1917, 1925, 1933, 1942, 1947 и 1962 годах), расположенного на Беломорском побережье предшествуют или совпадают с периодами всплесков естественного возобновления сосняков в бассейне Северского Донца (Вакуров, 1975; Салтыков, 2019). По данным А. Д. Вакурова исключительно засушливыми были тридцатые годы (1932, 1933, 1936–1939), когда лесные пожары в северных лесах проходили почти повсеместно. То же самое можно сказать о 1960, 1967, 1972 и 1973 годах. По сведениям автора, эти годы характеризовались высокой пожарной опасностью не только на Европейском Севере, но и в целом по стране (Вакуров, 1975). Начинаясь на юге равнины, атмосферные засухи неуклонно продвигаются на север, охватывая всё большие пространства, и несут за собой масштабные по площади лесные пожары. Последующая волна возобновления позволяет сосне восстановить популяционное пространство, избегая конкуренции со стороны темнохвойных и твердолиственных пород (Дмитриевский, 1928; Мелехов, 1944; Ткаченко, 1955; Врадий, 1961; Салтыков, 2019). В дальнейшем исследователи с определённой последовательностью уточняли и дополняли данные, касающиеся всплесков возобновления. Однако полученные результаты, в большинстве своём, служили отражением региональных особенностей формирования светлохвойных лесов (Вакуров, 1975). Более широкие обобщения, которые бы позволили выявить масштаб всплесков возобновления хвойных на пространстве равнины были затруднены, прежде всего, по причине отсутствия единства методического подхода. Кроме того, до настоящего времени существует крайне мало сведений относительно особенностей естественного возобновления в границах искусственно созданных насаждений, долевое участие которых в структуре лесов равнины, неуклонно возрастает (Ерохин, 2011; Грязькин и др., 2019; Салтыков, 2019).
Цель данного исследования – проверка рабочей гипотезы о синхронности всплеска возобновления сосняков естественного происхождения и культур P. sylvestris в границах южной покатости Русской равнины, а также о согласованности всплесков возобновления сосняков семиаридной зоны равнины и сосновых лесов с доминированием P. рallasiana в лесостепном предгорье Крымского полуострова.
ОБЪЕКТЫ И МЕТОДИКА ИССЛЕДОВАНИЙ
Исследования процессов естественного возобновления сосновых лесов на территории юго-западного сегмента Русской равнины были выполнены нами в течение 2003–2020 года. Отличительная черта западной половины равнины состоит в том, что её территория находится под отчётливо выраженным влиянием атлантических циклонов (Алисов, 1969; Швер, 1976; Мячкова, 1983). Периодическое увеличение количества поступающих осадков, обусловленное усилением циклонической деятельности и перераспределяющим эффектом поверхности равнины, отражается на биологическом разнообразии, продуктивности и устойчивости лесных насаждений. В связи с чем, объекты наблюдения разделены на три пространственно обособленных кластера: центральный, северный и южный.
Изначально изучение структурно—функциональных особенностей возобновления сосняков нами было намечено в границах центрального кластера – водосборного бассейна реки Северский Донец, площадь которого составляет около 100000 км2. В среднем течении реки бассейн Северского Донца разделён на лесостепную и степную зоны. Пристепные боры присутствуют на всем протяжении второй террасы, превращаясь в песчаную степь лишь вблизи его устья. Исторически сложившиеся особенности ведения лесного хозяйства на боровых террасах Северского Донца обусловили доминирование сосняков искусственного происхождения (Салтыков, 2019). За 12-летний период наблюдений (2003–2015 годы) с целью изучения особенностей естественного возобновления сосны обыкновенной заложено 572 пробных площади.
В 2015 году наши исследования были перенесены на территорию центральной части Русской равнины в зону хвойно-широколиственных лесов. Необходимость формирования северного кластера обусловливалась наличием сведений о синхронности всплесков возобновления не только в степной и лесостепной части Русской равнины, но и за их пределами (Пятницкий, 1964; Огиевский, 1966; Салтыков, 2019). Северная граница указанного кластера очерчена истоками реки Западная Двина. Южная граница опытных объектов совпадает с границами распространения Брянских лесных массивов, приуроченных к левому берегу реки Десны. В границах указанного кластера опытные объекты расположены на территории национальных парков «Смоленское Поозерье» и «Орловское Полесье», а также в биосферном заповеднике «Брянский лес». Полевые исследования были выполнены в течение трёх весенне-летних сезонов 20152017 годов. За указанный период времени заложено более 200 пробных площадей.
Итоги исследований 2003–2017 годов легли в основу выдвижения рабочей гипотезы о том, что всплески возобновления в условиях семиаридных зон будут согласованы по времени. В связи с чем, на южной границе Русской равнины был сформирован третий кластер с сетью опытных объектов, приуроченный к лесным экосистемам лесостепной предгорной части Крыма. С целью изучения пространственно-возрастной структуры подроста в горнолесном Крыму на протяжении 2017–2020 годах заложено 284 пробных площади.
При выполнении полевых исследований и формировании сети опытных объектов использована методика С. С. Пятницкого (1964). В рамках вариантов опыта, принятых к исследованию, предусмотрена четырёхкратная повторность. На каждой пробной площади выполнен перечёт подроста и установлены следующие показатели: количество и диаметр растений на высоте груди (см). В том случае, когда средняя высота подроста не превышала 1,3 м, выполнялись замеры диаметров на уровне шейки корня (см). Для тридцати случайно отобранных растений выполнены синхронные замеры: высоты (см), прироста верхушечной оси (см), размеров кроны во взаимно противоположных направлениях (см), установлен возраст (лет) и оценено жизненное состояние растения. Оценка особенностей возрастной структуры подроста и синхронности популяционных всплесков видов рода Pinus выполнена с привлечением методических подходов, разработанных Ю. А. Злобиным (1976, 2009). Полученные данные обработаны общепринятыми методами математической статистики.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Исследованиями установлено, что условием популяционного всплеска сосняков боровой террасы Северского Донца является совпадение повышенного уровня семеношения материнских насаждений с увеличением количества выпадающих осадков на фоне средней многолетней их величины (Дмитриевский, 1928; Врадий, 1961; Пятницкий, 1964; Салтыков, 2019).
Успешность возобновления пристепых боров, по мнению учёных, определяют весенне-летние осадки, обеспечивающие оптимальные условия для прорастания семян и укоренения всходов сосны (Дмитриевский, 1928; Краснов, 1950; Врадий, 1961; Пятницкий, 1964; Салтыков, 2019; Санников и др., 2019). Периоды с количеством осадков, заметно превышающих многолетнюю среднегодовую норму, закономерно повторяются, определяя временные границы возобновительного цикла. Указанное совпадение за 25-летний период наблюдения было характерным для 1995–1996, 2002–2003 и 2010–2011 года (рис. 1).
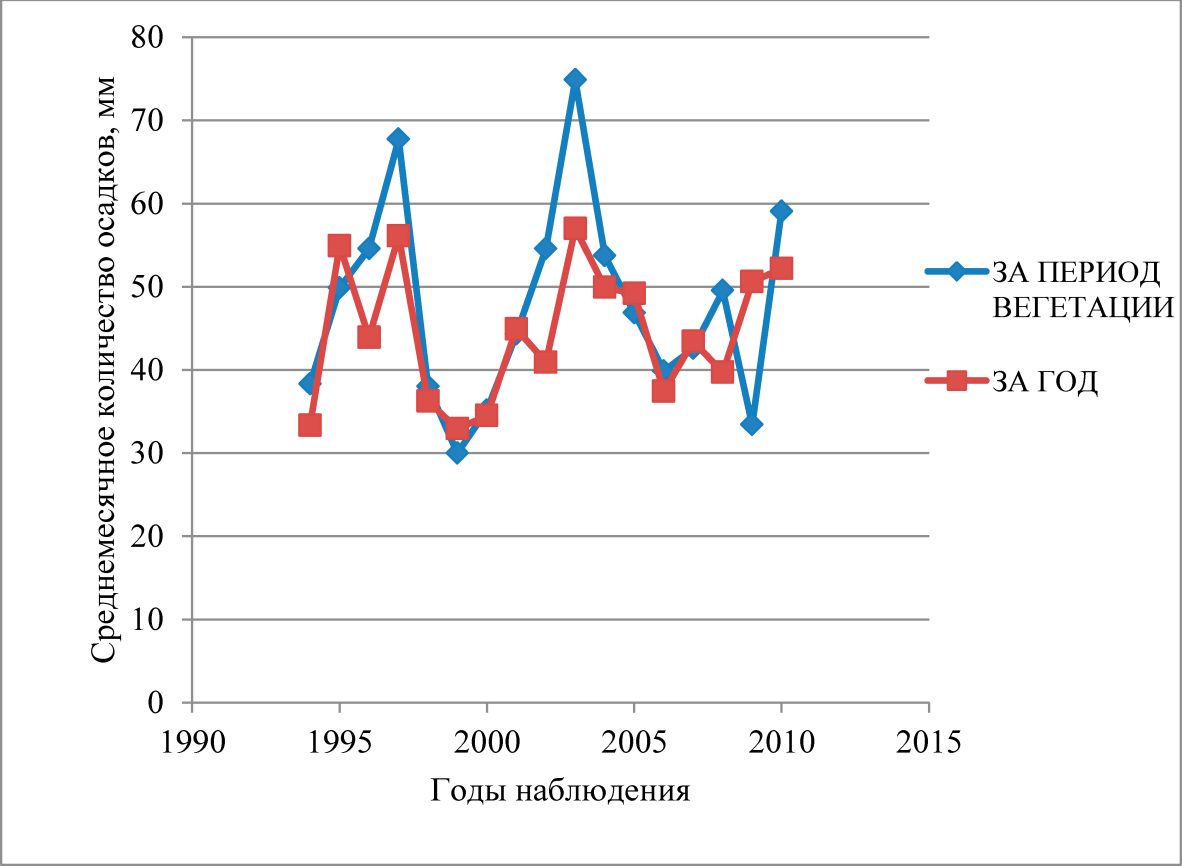
Рис. 1. Среднемесячное количество осадков по данным метеостанции Скрипаёвского учебно-опытного лесхоза Харьковского национального аграрного университета им. В. В. Докучаева, расположенной в среднем течении реки Северский Донец
Вне сомнения, популяционный всплеск, зафиксированный нами на боровых террасах Северского Донца в 2002–2003 годах (табл. 1) стал ответной реакцией лесных экосистем на оптимизацию гидротермического режима исследуемого региона. Появление всходов и самосева на боровой террасе наблюдалось повсеместно, однако последующая адаптация растений в условия существующих экологических ниш привнесла определённые коррективы в пространственно-возрастную структуру ценопопуляций.
Таблица 1
Фрагмент данных биометрической оценки подроста сосны обыкновенной в границах центрального кластера исследования
|
Лесохозяйственное (лесоохотничье) предприятие |
Шифр объекта |
Возраст, лет |
Доминанта возрастного спектра |
Происхождение материнского насаждения |
Экологические условия |
Высота, см |
|
Боровые террасы Северского Донца и его притоков |
||||||
| Волчанское |
52/07 Вс |
5,5±0,05 |
2002–2003 |
лесн. культ. |
с/х земли |
115,4±4,6 |
|
04/07 Вч |
4,1±0,01 |
2003–2002 |
лесн. культ. |
горельник |
13,4±0,3 |
|
| Балаклейское |
01/07 Бс |
5,1±0,03 |
2002–2003 |
природные |
горельник |
14,9±0,4 |
|
03/07 Бс |
6,0±0,00 |
2002 |
природные |
горельник |
14,9±0,4 |
|
| Змиевское |
01/10 Зз |
7,7±0,12 |
2002–2003 |
лесн. культ. |
с/х земли |
178,0±10,8 |
|
14/10 Зз |
7,4±0,29 |
2002–2003 |
лесн. культ. |
с/х земли |
184,3±26,7 |
|
| Скрипаевское |
72/09 Ш |
6.7±0,09 |
2002–2003 |
лесн. культ. |
с/х земли |
158,5±4,2 |
|
72/4 Сл |
7,2±0,08 |
2002–2003 |
природные |
горельник |
144,9±6,9 |
|
| Изюмское |
03/08 Ип |
5,9±0,11 |
2002–2003 |
природные |
горельник |
65,1±4,1 |
|
04/08 Ип |
5,8±0,06 |
2002–2003 |
природные |
горельник |
50,2±2,7 |
|
| Краснолиманское |
28/10 Кя |
7,9±0,05 |
2002–2003 |
лесн. культ. |
горельник |
207,2±7,0 |
|
04/10 Кд |
7,9±0,07 |
2002–2003 |
лесн. культ. |
горельник |
73,5±3,7 |
|
| Кременское |
120/06 К |
5,0±0,02 |
2002–2003 |
природные |
горельник |
50,5±2,2 |
|
03/11 Кб |
9,0±0,00 |
2002–2003 |
лесн. культ. |
с/х земли |
156,8±5,0 |
|
| Северодонецкое |
01/11 Сб |
9,1±0,07 |
2002–2003 |
лесн. культ. |
с/х земли |
222,9±12,2 |
|
11/13 Сб |
9,9±0,03 |
2002–2003 |
лесн. культ. |
горельник |
139,3±7,1 |
|
| Станично-луганское |
03/09 Сл |
6,2±0,13 |
2002–2003 |
природные |
горельник |
57,9±4,4 |
|
05/09 Сл |
6,9±0,05 |
2002–2003 |
природные |
горельник |
104,2±7,3 |
|
|
Боровые террасы Днепра и его притоков |
||||||
| Близнюковское |
12/09 Бл |
6,8±0,10 |
2002–2003 |
лесн. культ. |
горельник |
106,7±6,1 |
|
11/09 Бл |
6,8±0,07 |
2002–2003 |
лесн. культ. |
горельник |
131,1±5,3 |
|
| Красноградское |
04/10 Кр |
7,3±0,16 |
2002–2003 |
лесн. культ. |
с/х земли |
221,4±16,6 |
|
08/10 Кн |
7,1±0,17 |
2002–2003 |
лесн. культ. |
с/х земли |
180,4±15,0 |
|
| Новосанжарское |
01/12 Н |
9,7±0,05 |
2002–2003 |
лесн. культ. |
горельник |
151,3±5,6 |
|
02/12 Н |
9,9±0,02 |
2002–2003 |
лесн. культ. |
горельник |
154,9±5,0 |
|
| Семеновское |
06/11 С |
6,8±0,07 |
2003–2004 |
не установл. |
с/х земли |
222,8±8,3 |
|
07/11 С |
6,5±0,08 |
2003–2004 |
не установл. |
с/х земли |
199,9±7,3 |
|
С течением времени жизнеспособный подрост сосны был зафиксирован нами лишь по внешнему линейному контуру лесных массивов и в «окнах» полога материнских насаждений, в то время как под пологом леса наблюдался процесс затухания волны возобновления. Завершение популяционного всплеска сопровождалось расслоением единого поля возобновления в сеть пространственно разобщённых объектов с динамично меняющейся структурой подроста. Обязательным условием и характерной чертой процесса является выраженная приуроченность самосева и подроста к условиям пирогенного ряда, либо землям, выведенным из-под сельскохозяйственного пользования (Дмитриевский, 1928; Салтыков, 2019).
Успешная реализация репродуктивного потенциала насаждений в самосев и подрост во многом определена ёмкостью ниши возобновления, её соответствием биоэкологическим свойствам растений. В каждом конкретном случае структура ценопопуляции или субценопопуляционного фрагмента и ниша возобновления, являясь комплементарными парами, определяют жизненное состояние подроста и динамику его роста и развития. Примером авторегуляции структуры ценопопуляции в условиях существующей экологической ниши является дрейф доминанты возрастного спектра подроста в границах периода популяционного всплеска (табл. 2).
Таблица 2
Варьирование доминирующего поколения подроста сосны в период популяционного всплеска
|
Шифр пробы |
Доминирующее поколение, год |
Дата наблюдения, год |
Вид пожара |
Тип леса |
Количество тыс. шт./га |
|
04/07 Вч |
2003 |
весна 2007 |
низовой |
В2–дС |
122,1 |
|
01/07 Бс |
2002 |
весна 2007 |
низовой |
В2–дС |
226,8 |
|
28/10 Кя |
2002 |
весна 2010 |
переходной |
А2–С |
22,4 |
В приведённом примере смещение доминанты возрастного спектра было обусловлено следствием прохождения лесных пожаров. В первом случае реализация репродуктивного потенциала насаждения состоялась сразу после низового пожара весной 2003 года на этапе завершения популяционного всплеска, во втором и третьем в 2002 году. Аналогичное смещение доминанты возрастного спектра возможно в противоположную сторону, когда в границах ценопопуляции доминируют растения появившееся в 2001 году (Салтыков, 2019). То есть на общем фоне всплеска возобновления и преобладания подроста, который появился в 2002 году, ожидаемо присутствие ценопопуляций со смещением доминанты спектра на один год (2002±1 год). В качестве примера доминирования поколения 2002 года, нами приведены иллюстрации возрастных спектров (рис. 2) сосны обыкновенной по данным пробных площадей: а) 01/07 Бс и б) 28/10 Кя. Для пробы 04/07 Вч характерно абсолютное доминирование поколения 2003 года, обусловленное массовым появлением всходов и самосева сосны сразу после прохождения низового пожара.
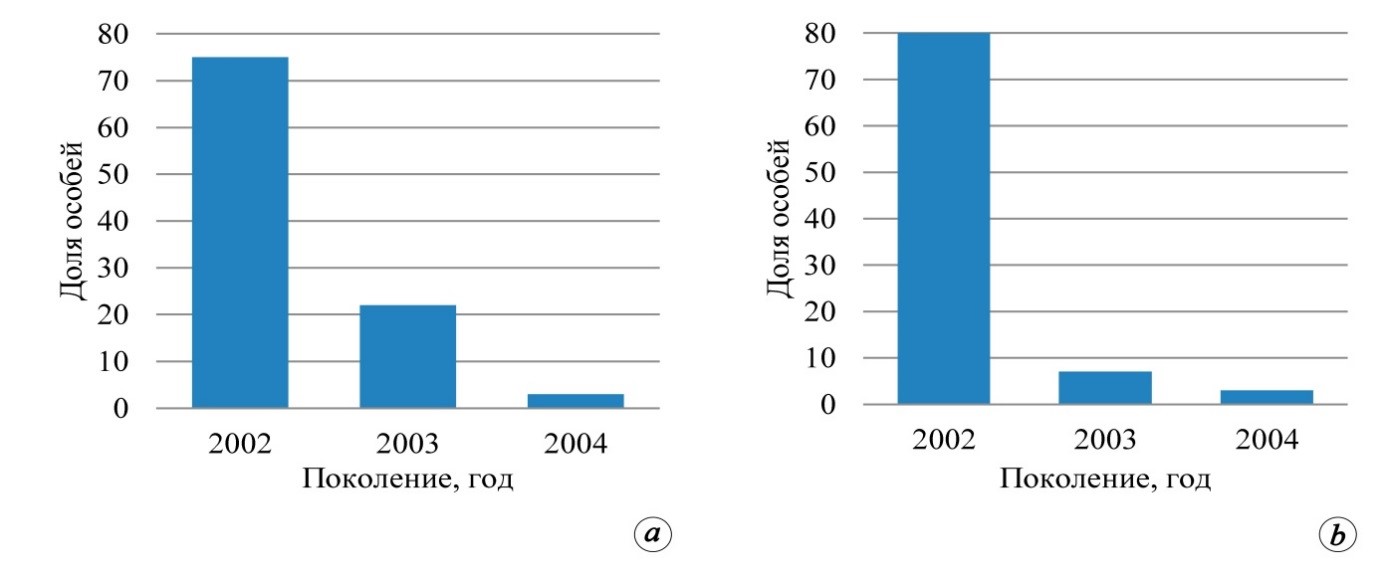
Рис. 2. Возрастные спектры подроста сосны обыкновенной на пробных площадях, заложенных на горельниках: 01/07 Бс (а) и 28/10 Кя (б)
Более заметные изменения структуры возрастного спектра составляют, как правило, исключение и, вероятны, для очень сухих и сухих боровых условий поскольку в условиях крайне бедных местообитаний длительный период восстановления напочвенного покрова отражается на процессах возобновления и, соответственно, на структуре возрастных спектров, размывая их границы во времени. Таким образом, авторегуляция структуры ценопопуляции и, как частный пример, дрейф доминанты возрастного спектра обусловлены наличием и ёмкостью ниш возобновления, в той или иной мере отвечающих за успешность реализации репродуктивного потенциала сосняков в категорию самосева и подроста. Варьирование пространственно-возрастной структуры ценопопуляций подроста в соответствии с разнообразием экологических условий служит подтверждением позитивной тенденции процесса возобновления в пространстве боровой террасы.
Исследованиями установлено, что жизнеспособный подрост сосны присутствует на территории всех лесохозяйственных предприятий расположенных как в лесостепной, так и в степной зоне Северского Донца (табл. 1). Синхронность всплеска возобновления в равной мере свойственна коренным древостоям сосны обыкновенной и искусственно созданным насаждениям (табл. 1), а присутствие жизнеспособных ценопопуляций подроста в одной из географических точек бассейна служит основанием для утверждения, что с большой долей вероятности подрост этой же генерации можно встретить в любой другой точке боровой террасы. Наряду с жизнеспособными ценопопуляциями подроста, которые сформировались в период 2002±1 году в лесных массивах присутствуют молодняки сосны с доминантой 1995–1996 годов. Указанная категория молодняков, как правило, приурочена к различного рода разрывам в пологе материнских насаждений или же внешним линейным контурам лесных массивов. Краткие сведения о наличии молодняков с доминантой возрастного спектра 1995–1996 годов приведены в таблице 3.
Таблица 3
Доминирующие поколения подроста сосны обыкновенной в границах центрального кластера исследований
|
Лесохозяйственное (лесоохотничье) предприятие |
Доминирующее поколение |
Экологические условия |
Тип ценопопуляции подроста |
| Волчанское |
1995–1996 |
пирогенный ряд | устойчивый |
|
2002–2003 |
земли с.-х. назначения | процветающий | |
| Змиевское |
1993–1995 |
земли с.-х. назначения | процветающий |
|
2002–2003 |
земли с.-х. назначения | процветающий | |
| Скрипаёвское |
1993–1995 |
пирогенный ряд | устойчивый |
|
2002–2003 |
пирогенный ряд | процветающий | |
| Чугуево-Бабчанское |
2002–2003 |
пирогенный ряд | процветающий |
|
1995–1996 |
пирогенный ряд | процветающий | |
| Балаклейское |
1995–1996 |
пирогенный ряд | депрессивный |
|
2002–2003 |
пирогенный ряд | депрессивный | |
| Краснолиманское |
1995–1996 |
пирогенный ряд | процветающий |
|
2002–2003 |
пирогенный ряд | процветающий | |
| Кременское |
1994–1996 |
пирогенный ряд | процветающий |
|
2002–2003 |
земли с.-х. назначения | процветающий | |
| Близнюковское |
1995–1996 |
пирогенный ряд | процветающий |
|
2002–2003 |
земли с.-х. назначения | процветающий | |
| Красноградское |
1995–1996 |
пирогенный ряд | процветающий |
|
2002–2003 |
земли с.-х. назначения | процветающий | |
| Северодонецкое |
1995–1996 |
пирогенный ряд | процветающий |
|
2002–2003 |
земли с.-х. назначения | процветающий | |
| Станично-Луганское |
1995–1996 |
пирогенный ряд | процветающий |
|
2002–2003 |
пирогенный ряд | процветающий |
Молодняки сосны адаптированы в существующие ниши возобновления, подтверждением чему является их жизненное состояние (табл. 3). Сочетание в пространстве лесных массивов, по меньшей мере, двух генераций растительных группировок сосны позволяет выдвинуть предположение о том, что по мере формирования пространства возобновления, периодические повторяющиеся всплески направлены на восстановление утраченного жизненного пространства сосняков. Каждый последующий всплеск естественного возобновления является основой совершенствования структуры и обеспечения устойчивости популяции P. sylvestris. Пульсирующий характер естественного возобновления пристепных боров обусловлен климатическими особенностями семиаридной зоны Русской равнины, прежде всего, циклоничностью Атлантики (Алисов, 1969; Мячкова, 1983; Салтыков, 2019).
Вероятно, в регионах, где отсутствует лимитирующее влияние влаги, подобная закономерность периодичности популяционных всплесков исключена. Типичным примером является зона хвойно-широколиственных лесов Русской равнины, где количество осадков равно или заметно превышает испарение (Алисов, 1969; Мячкова, 1983; Салтыков, 2019). С целью проверки данного предположения изучение пространственно-возрастной структуры подроста и молодняков сосны были продолжены в границах северного кластера. Результаты исследований показали, что на территории национальных парков «Смоленское Поозерье», «Орловское Полесье» и биосферного заповедника «Брянский лес» присутствуют ценопопуляции подроста и молодняки сосны со схожими доминантами возрастных спектров. Так, например, доминирующее поколение молодняков сосны 1995–1996 годов зафиксировано нами на территории биосферного заповедника «Брянский лес» и национального парка «Смоленское Поозерье» (табл. 4).
Таблица 4
Доминирующие поколения подроста сосны обыкновенной в границах северного кластера исследования
|
Шифр пробы |
Возраст, лет |
Доминирующее поколение, год |
Высота, м |
Количество тыс. шт./га |
Тип ценопопуляции |
|
Биосферный заповедник «Брянский лес» |
|||||
| 25/16 Бл |
19,7±0,12 |
1995–1996 |
6,8±0,56 |
6,9 |
процветающий |
| 29/16 Бл |
19,7±0,10 |
1995–1996 |
6,7±0,22 |
15,3 |
процветающий |
|
Национальный парк «Смоленское Поозерье» |
|||||
|
21/16 Сп |
19,8±0,23 |
1995–1996 |
10,5±0,09 |
4,5 |
процветающий |
|
25/16Сп |
19,4±0,18 |
1995–1996 |
10,6±0,10 |
4,0 |
процветающий |
Молодняки сосны с указанной возрастной доминантой имели повсеместное распространение на боровой террасе Северского Донца. Также в границах зоны хвойно-широколиственных лесов нами были зафиксированы молодняки сосны, формирование которых следует отнести к 2002–2003 годам. Достаточно широко представлена указанная категория молодняков на территории национального парка «Смоленское Поозерье». Объединяет указанные объекты не только доминирование поколений 2002–2003 годов, но и выраженная их принадлежность к определённым экологическим условиям. Так, в большинстве своём массивы таких сосновых молодняков расположены либо на землях, выведенных из-под сельскохозяйственного пользования, или же размещены в границах пирогенного ряда (табл. 5).
В качестве иллюстрации, позволяющей продемонстрировать доминирование генерации сосны, появившейся в 2002±1 год, ниже приведены возрастные спектры ценопопуляций подроста на опытных объектах (рис. 3).
Данные выполненных нами исследований позволяют сделать предположение о том, что ценопопуляции подроста сосны с возрастной доминантой 2002±1 года с большой долей вероятности ожидаемы на территории западного сегмента зоны хвойно-широколиственных лесов. Кроме указанной категории молодняков сосны на территории национальных парков «Смоленское Поозерье», «Орловское Полесье» и биосферного заповедника «Брянский лес»
Таблица 5
Доминирующие поколения подроста сосны обыкновенной в границах северного кластера исследований (национальный парк «Смоленское Поозерье»)
|
Шифр пробы |
Возраст, лет |
Доминирующее поколение, год |
Высота, м |
Количество тыс. шт./га |
Тип ценопопуляции |
|
1/15 Сп |
13,4±0,11 |
2002–2003 |
5,9±0,33 |
5,6 |
процветающий |
|
5/15 Сп |
13,7±0,15 |
2002–2003 |
5,8±0,27 |
7,5 |
процветающий |
|
9/15 Сп |
13,5±0,11 |
2002–2003 |
5,7±0,25 |
6,0 |
процветающий |
|
13/15 Сп |
12,6±0,14 |
2002–2003 |
4,1±0,26 |
5,4 |
процветающий |
|
17/15 Сп |
13,1±0,09 |
2002–2003 |
5,1±0,22 |
3,8 |
процветающий |
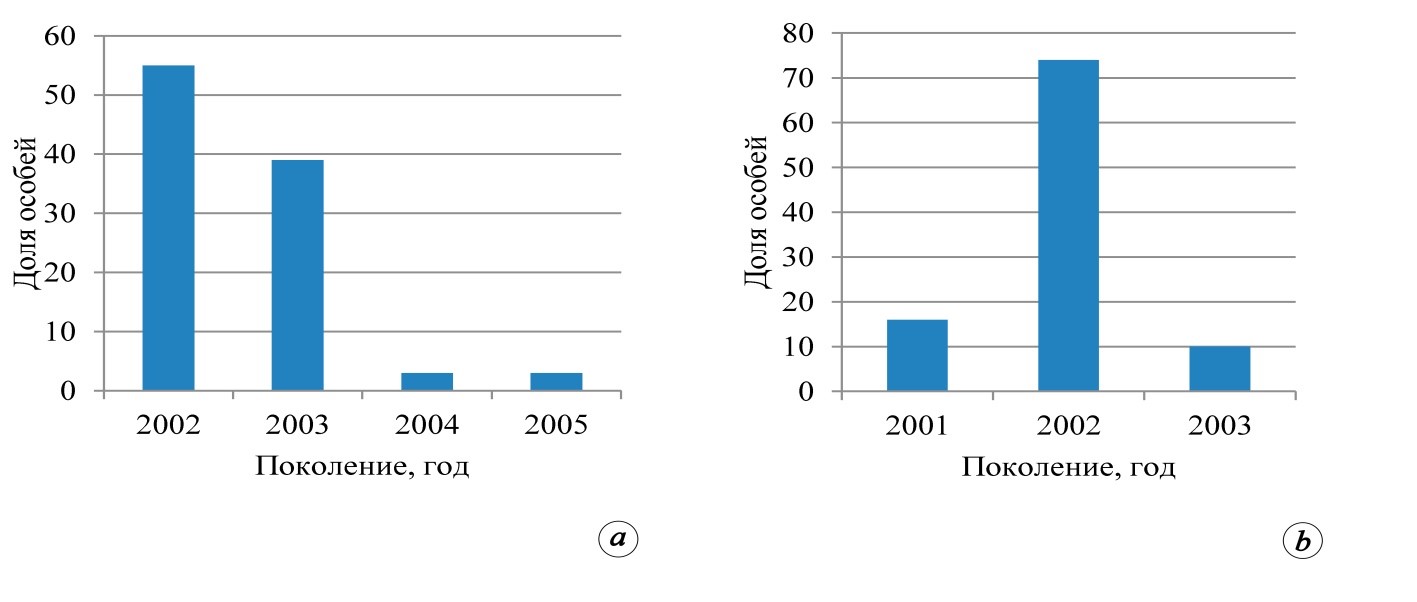
Рис. 3. Возрастные спектры подроста Pinus sylvestris на пробных площадях:
13/15 Сп (а) и 17/15 Сп (б)
зафиксированы ценопопуляции подроста с доминантой возрастного спектра в 2007–2008 годов. Эта же возрастная категория подроста осенью 2014 года нами была выявлена на боровой террасе Северского Донца. Синхронность всплесков возобновления рода Pinus во времени в бассейне Северского Донца и на объектах, расположенных в зоне хвойно-широколиственных не случайна. Одна из наиболее вероятных причин, по мнению исследователей, состоит в климатической однородности исследуемых территорий. «Совпадение между составляющими теплового баланса, однородности циркуляционных процессов, особенности годового хода основных показателей климата заставляет объединить лесостепь ЕТС и юг лесной области в одну климатическую область» (Мячкова, 1983). Очевидно, что и ответная реакция лесных экосистем на влияние комплекса абиотических факторов будет близкой, что и прослеживается на всплесках возобновления сосняков в регионе исследования. Периодичность и в тоже время постоянство популяционных потоков P. sylvestris в границах опытных объектов позволяет верифицировать предположение о синхронности популяционных всплесков сосны обыкновенной на территории южной покатости Русской равнины, как в лесостепной и степной зонах, так и за их пределами, прежде всего, в границах западного сегмента зоны хвойно-широколиственных лесов.
Не вызывающий сомнений пульсирующий характер процессов естественного возобновления в условиях семиаридной зоны равнины послужил основанием для изучения пространственно-возрастной структуры подроста и молодняков сосны в условиях лесостепного предгорья Крыма. За основу исследований была принята рабочая гипотеза о том, что всплески естественного возобновления сосны крымской будут синхронизированы или же близки по времени с установленными нами ранее датами популяционных всплесков
Таблица 6
Фрагмент оценки возрастной структуры подроста и молодняков сосны крымской на объектах исследования
|
Объект |
Возраст, лет |
Доминирующее поколение, год |
Высота, м |
Тип ценопопуляции |
|
Белогорское лесничество |
||||
|
40/44 Бг |
20,9±0,27 |
1996‒1997 |
3,5±0,16 |
процветающий |
|
Новокленовское лесничество |
||||
|
19/19 Нк |
17,0±0,21 |
2002–2003 |
5,2±0,41 |
процветающий |
|
22/19 Нк |
16,2±0,23 |
2002–2003 |
1,6±0,01 |
устойчивый |
|
Симферопольское лесничество |
||||
|
83/18 Сф |
16,7±0,76 |
2001–2003 |
3,9±0,25 |
процветающий |
|
Солнечногорское лесничество |
||||
|
251/17 Сг |
16,9±0,18 |
2002–2003 |
3,9±0,19 |
процветающий |
|
256/17 Сг |
15,7±0,18 |
2002–2003 |
3,7±0,18 |
процветающий |
|
261/17 Сг |
15,4±0,33 |
2002–2003 |
3,5±0,17 |
процветающий |
для лесостепной и степной зоны равнинной части Восточно-Европейской равнины. В связи с этим в границах предгорной части Крыма нами была заложена сеть опытных объектов. Краткие итоги выполненных наблюдений по оценке возрастной структуры и состоянию подроста и молодняков сосны отражены таблице 6.
На территории Белогорского, Новокленовского, Симферопольского и Солнечногорского лесничеств было установлено присутствие молодняков P. рallasiana с доминантами возрастных спектров 2002±1, 1996±1 годов (рис. 4). В Белогорском лесничестве подрост и молодняки сосны естественного происхождения приурочены к противопожарным разрывам. В Новоклёновском участком лесничестве сосновые молодняки приурочены к землям, выведенным из-под сельскохозяйственного пользования. Также указанная категория молодняков естественного происхождения примыкает к внешним контурам и распространена в окнах полога материнских насаждений. При этом разница жизненного состояния молодняков P. рallasiana значительна. Так, например, в границах существующих разрывов в пологе насаждений растения отстали в росте и развитии, угнетены, являются ближайшими кандидатами на отпад. Под пологом насаждений сохранились лишь единичные нежизнеспособные, случайно размещённые в пространстве материнских насаждений особи подроста.
По внешним контурам и за пределами влияния материнских насаждений молодняки сосны отличаются хорошим ростом и развитием. В Солнечногорском лесничестве молодняки сосны, сформировавшиеся в 2002–2003 годах своим происхождением обязаны плодоношению культур сосны крымской, созданных на террасах горнолесного Крыма во второй половине прошлого столетия (Ткач и др., 2013). В настоящее время они успешно освоили прилегающее к лесным полосам пространство, удаление от стен леса естественным образом сформированных молодняков достигает 100–200 м и более. Перспектива их дальнейшего роста и развития не вызывает сомнений. Можно предполагать, что сосна крымская успешно восстанавливает ранее утраченное ранее жизненное пространство.
Опираясь на полученные данные, можно сказать, что в границах южного кластера исследований отчётливо выделяются ценопопуляции подроста и молодняков сосны крымской с возрастной доминантой 1996±1 года, 2002±1 года. Существует вероятность того, что пики возобновления в указанные временные промежутки были связаны с изменением гидротермического режима лесных экосистем. Так, например, за 20-летний период (1986‒2005 годы) годы 1997 и 2002-й отличались тем, что количество осадков выпало значительно
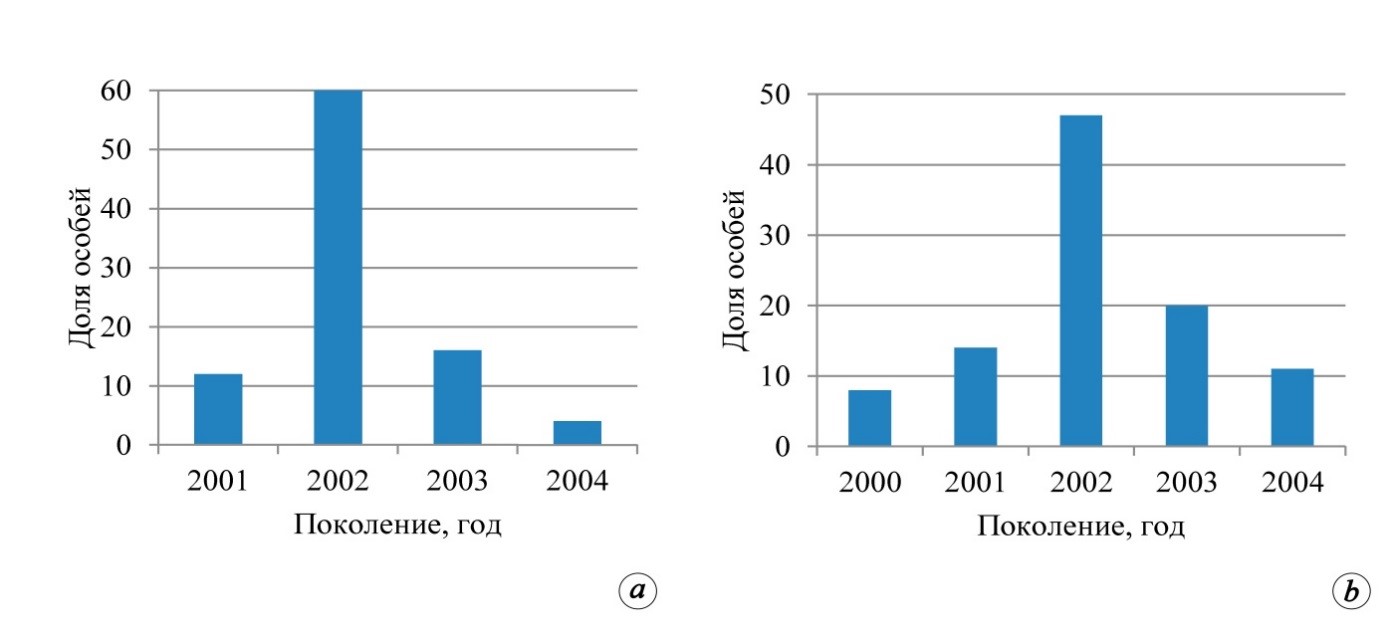
Рис. 4. Возрастные спектры подроста сосны крымской с доминированием поколения 2002 года на пробных площадях: 19/19 Нк (а) и 25/17 Сг (б)
больше многолетней среднегодовой нормы. При этом в 1997 году отмечено абсолютное увеличение осадков, в то время как в 2002 году превышение многолетней нормы было характерно, прежде всего, для вегетационного периода (Агрокліматичний довідник…, 2011). Очевидно, что подобное совпадение и следующий за ним всплеск возобновления P. рallasiana синхронизирован с аналогичными процессами лесостепной и степной зоны Русской равнины. Причиной периодичности всплесков являются глобальные природные процессы, обусловленные активностью атлантических циклонов. Периоды со снижением указанного процесса приводят к формированию своеобразных экологических барьеров, что определяет длительность цикла возобновления и наличие временных разрывов в границах возрастных спектров ценопопуляций.
ЗАКЛЮЧЕНИЕ
Пульсирующий характер естественного возобновления Pinus sylvestris предопределён климатическими особенностями семиаридной зоны Русской равнины, прежде всего, динамикой прихода осадков. Популяционные всплески согласованы с глобальными природными процессами, массовое появление всходов и самосева сосны обыкновенной в границах лесостепной и степной зоны является следствием оптимизации гидротермического режима лесных экосистем во время активизации циклонической деятельности Атлантики. Засушливые периоды со снижением количества выпадающих осадков приводят к формированию экологических барьеров, определяющих длительность цикла возобновления и дискретность возрастных спектров ценопопуляций во времени.
Шлейф синхронности популяционных всплесков представителей рода Pinus прослеживается на обширных пространствах равнины и в её приграничных зонах, не ограничиваясь только границами степной и лесостепной зоны. Так, например, жизнеспособный подрост и молодняки сосны обыкновенной и крымской с доминантой 2002±1 и 1996±1 года кроме лесостепной и степной зоны равнины зафиксирован нами в зоне хвойно-широколиственных лесов и в горно-лесном Крыму и, прежде всего, в лесостепном предгорье. Вероятной причиной согласованности всплесков возобновления во времени на территории юго-западного сегмента Русской равнины является климатическая однородность исследуемых территорий.
Периодические повторяющиеся всплески естественного возобновления направлены на восстановление утраченного популяционного пространства сосняков. Синхронность всплеска возобновления в равной мере свойственна коренным древостоям сосны обыкновенной и искусственно созданным насаждениям. Пространственно-возрастная структура ценопопуляций подроста и молодняков сосны обыкновенной и крымской и её варьирование в соответствии с разнообразием экологических условий позволяет каждой последующей волне возобновления освоить все потенциально возможные экологические ниши и в тоже время служит подтверждением позитивной тенденции процесса возобновления в пространстве сосновых лесов равнины и прилегающих территорий.
REFERENCES
Агрокліматичний довідник по Автономній республіці Крим (1986– 2005 рр.) / Ред. О. І. Прудко, Т. І. Адаменко. – Сімферополь: «Таврида», 2011. – 343 с.
Алисов Б. П. Климат СССР. – Москва, «Высшая школа», 1969. – 131 с.
Вакуров А. Д. Лесные пожары на Севере. – Москва: Наука, 1975. – 100 с.
Врадий Н. И. Пристепные боры Украины и способы создания в них лесных культур: дис. … канд. с-х. наук. – Харьков, 1961. – 365 с.
Высоцкий Г. Н. О боровых типах Чугуево-Бабчанского лесничества вблизи Харькова на Северном Донце. Очерки по фитоценологии. – Москва, 1929 – С. 7–15.
Грязькин А. В., Беляева Н. В., Шахов А. Г., Нгуен Ван Зинь Естественная смена ели сосной на участках лесных культур // Лесотехнический журнал, 2019. Т. 9, № 1 (33). С. 5460.
Дмитриевский П. И. К вопросу о возобновлении сосновых лесов естественным подростом // Вести ХСХИ, 1928, № 10. – С. 1–19.
Ерохин А. В. Естественное возобновление в культурах сосны // Актуальные проблемы лесного комплекса. – 2011. Вып. 30. С. 1315.
Злобин Ю. А. Оценка качества ценопопуляций подроста древесных пород // Лесоведение, 1976. – № 6. – С. 72–79.
Злобин Ю. А. Популяционная экология растений: современное состояние, точки роста. – Сумы: Университетская книга, 2009. – 263 с.
Коба В. П. Особенности восстановления древостоев сосны крымской в постпирогенный период // Экосистемы, 2017. – Вып. 11. – С. 10–13.
Краснов М. А. Естественное возобновление сосны в связи с рубками и пожарами // Бузулукский бор. – Москва–Ленинград: Гослесбумиздат, 1950. – Т. II. – C. 3–97.
Мелехов И. С. О теоретических основах лесной пирологии. – Архангельск: Архангельский лесотехнический институт, 1944. – 20 с.
Мячкова Н. А. Климат СССР. – Москва: Издательство МГУ, 1983. – 192 с.
Огиевский В. Д. Избранные труды. – Москва: Лесная пром-сть, 1966. – 356 с.
Одум Ю. Экология. В 2-х тт. Москва: Мир. 1986. Т.1 328 с., Т. 2376 с.
Переведенцев Ю. П., Верещагин М. А., Шанталинский К. М., Наумов Э. П., Хабутдинов Ю. Г. Изменения климатических условий и ресурсов Среднего Поволжья: учебное пособие по региональной климатологии / Ред. Э. П. Наумов. – Казань: Центр инновационных технологий, 2011. – 296 с.
Поликарпов Н. П. Эколого-географические закономерности естественного лесовозобновления // Лесное хозяйство. – 1978. – № 3. – С. 60 – 63.
Пятницкий С. С. Лесовозобновление в условиях левобережной Лесостепи УССР. Т. ХLV. – Киев, 1964. С. 3–23.
Пятницкий С. С. Методика исследований естественного семенного возобновления в лесах левобережной Лесостепи Украины. – Харьков, 1959. – 26 с.
Ткач В. П., Агапонов Н. Н. Защитные леса и лесные мелиорации горного Крыма. – Харьков: Планета-принт, 2013. – 320 с.
Риклефс Р. Основы общей экологии / [Ред. Н. Н. Карташев]. – Москва: Мир, 1979. – 424 с.
Салтыков А. Н. Структурно-функциональные особенности естественного возобновления придонецких боров. – Симферополь: И Т «Ариал». – 2019. – 361 с.
Санников С. Н., Санникова Н. С., Кочубей А. А., Петрова И. В. Естественное возобновление сосны на гарях в лесостепи Западной Сибири // Сибирский лесной журнал. – 2019. № 5. С. 2229.
Синицын Е. Н. Естественное возобновление сосняков Усманского и Хреновского боров. – Воронеж: ВГПУ, 2008. – 307 с.
Ткаченко М. Е. Общее лесоводство / [Ред. И. С. Мелехов. – Москва; Ленинград: Гослесбумиздат, 1955. – 600 с.
Тюрин А. В. Основы хозяйства в сосновых лесах. – Москва; Ленинград: Гослесбумиздат, 1952. – 112 с.
Флора европейской части СССР / [Ред. А. Н. Федоров]. – Ленинград: Наука, 1974. – Т. I. – 404 с.
