
STABILIZATION AND DECOMPOSITION RATE OF STANDARD MATERIAL IN HERBACEOUS COMMUNITIES OF THE FOREST ZONE
JOURNAL: «EKOSISTEMY», Issue 25, 2021
Publication text (PDF): Download
UDK:582.099-044.967:631.415(470.311)
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:Elumeeva T. G., Cherednichenko O. V., Gavrilova T. M. Lomonosov Moscow State University, Moscow, Russia
TYPE: Article
DOI: https://doi.org/10.37279/2414-4738-2021-25-12-21
PAGES: from 12 to 21
STATUS: Published
LANGUAGE:Russian
KEYWORDS:soil reaction, Tea Bag Index (TBI), stabilization factor, decomposition constant, herb vegetation, Tver region, Moscow region.
ABSTRACT (ENGLISH):
The rate of plant litter decomposition depends on two simultaneous processes: the decomposition and stabilization of organic matter. To compare the rates of these processes in various ecosystems now a Tea Bag Index (TBI) is widely used. The samples of standard material (green tea and rooibos) were buried into soil at the depth of 8 cm in four herbaceous plant communities of Central Forest State Nature Biosphere Reserve (CFSNBR) and seven communities of the Moscow river floodplain near the MSU Zvenigorod biological station (ZBS). Considering the mass losses of samples during the incubation time we calculated the values of decomposition constant and stabilization factor according to the protocol by J. A. Keuskamp et al. (2013) and estimated the links of these parameters with Ellenberg’s ecological indicator values. The stabilization factor values ranged from 0.122 g/g in Filipendula community of CFSNBR to 0.379 g/g at the natural levee of the Moscow river, and the decomposition constant values ranged from 0.0089 g/g/day in Filipendula community of CFSNBR to 0.0308 g/g/day at the slope of the high floodplain. The mean values of these parameters were significantly positively correlated. The decomposition rate was lower in the communities of CFSNBR and was positively correlated with the soil pH Ellenberg’s indicator values. The stabilization factor values did not depend on the region and decreased with the increase in soil moisture and nitrogen. The mainland meadows of CFSNBR and the meadows of the high floodplain of the Moscow river, belonging to Cynosurion cristati alliance, had the most similar parameters of decomposition.
ВВЕДЕНИЕ
Процессы разложения растительных остатков – важная часть круговорота веществ в экосистемах. Их скорость складывается из двух параметров: стабилизации органического вещества, например, в результате физической изоляции от сообщества разлагающих его микроорганизмов или образования органо-минеральных соединений, и собственно разложения с выделением углекислого газа. В свою очередь, стабилизация и разложение зависят от факторов среды, влияющих на деятельность микроорганизмов: климата, физических и химических свойств почвы, а также от качества самих отмерших растительных остатков (Prescott, 2010; Stockmann et al., 2013; Rowley et al. 2018). Функциональные признаки органов растений, например, соотношение азота и углерода в тканях, обусловливают их быстрое или медленное разложение после отмирания (Freschet et al., 2012), в связи с чем, могут влиять на его скорость сильнее, чем факторы экотопа (Fanin et al., 2019). В связи с этим Й. А. Кеускампом с соавторами (Keuskamp et al., 2013) был предложен метод для оценки фактора стабилизации (stabilization factor, S) и константы скорости разложения лабильной фракции (decomposition constant, k) на основе потери массы образцами легко доступного стандартного материала – пакетированного зеленого чая (Camellia sinensis, Theaceae) с легко разлагающимися листьями и ройбоса (Aspalthus linearis, Fabaceae) с относительно медленно разлагающимися листьями, так называемый индекс «чайных пакетиков» (Tea bag index, TBI). Фактор стабилизации показывает, какая часть лабильной фракции органического вещества переходит в трудно разлагаемые соединения, а константа скорости разложения – насколько быстро идет его разложение с учетом стабилизации. В настоящее время этот метод довольно часто используется для сравнения различных экотопов по параметрам разложения (Fujii et al., 2017; Whigham et al., 2017; Elumeeva et al., 2018; McDonald et al., 2018; Petraglia et al., 2019; Saint-Laurent, Arsenault-Boucher, 2020; Fanin et al., 2020). В России работы, проведенные по данной методике, единичны (Elumeeva et al., 2018).
В данной статье приведены результаты двух опытов с использованием TBI для травяных сообществ Московской области (Звенигородская биостанция МГУ) и Тверской области (Центрально-Лесной государственный природный биосферный заповедник). Целью нашей работы была оценка фактора стабилизации (STBI) и константы разложения (kTBI) в различных травяных сообществах и их связей с характеристиками экотопа, а также проверка гипотезы о том, что относящиеся к близким синтаксонам сообщества обладают сходными параметрами разложения.
МАТЕРИАЛ И МЕТОДЫ
Район исследования. Объектами исследования были выбраны семь травяных сообществ поймы реки Москвы на территории Звенигородской биостанции МГУ имени С. Н. Скадовского (ЗБС, Одинцовский район Московской области, 55°42′ с. ш., 036°44′ в. д.) и четыре травяных сообщества Центрально-Лесного государственного природного биосферного заповедника и его охранной зоны (ЦЛГПБЗ, Нелидовский район Тверской области, 56°26’–56°39′ с. ш. 032°29’–033°01′ в. д.). Краткая характеристика сообществ приведена в таблице 1.
ЗБС располагается в центре Русской равнины на правом берегу реки Москвы, территория включает узкую полосу современной поймы, древние террасы и водораздельную часть (Копцик, Рыжова, 2011; Александровский, Панин, 2013). Среднегодовая температура составляет 5,6 °C, сумма осадков – 634,7 мм (Погода и климат). Основную часть поймы реки Москвы на ее территории занимает доголоценовая пойма, которая приподнята на 5–7 м над рекой и нерегулярно заливалась в голоцене, а после постройки Можайского водохранилища в XX веке не заливалась. Современная пойма, сформировавшаяся в голоцене, слабо дифференцирована, на некоторых участках в ней выделяются прирусловой вал и центральная пойма, которые заливаются ежегодно (Чередниченко, Первова, 2016). Заливаемые участки заняты высокотравной нитрофильной растительностью класса Epilobietea angustifolii Tx. et Preising ex von Rochow 1951, а плоские участки высокой поймы и склон от высокой поймы к низкой – типичными лугами класса Molinio-Arrhenatheretea R. Tx. 1937 (Чередниченко, Ямалов, 2011). В понижениях формируются низинные болота и таволговые сообщества.
ЦЛГПБЗ располагается на юго-западе Валдайской возвышенности на Каспийско-Балтийском водоразделе Русской равнины. Климат гумидный континентальный (Минаева, Шапошников, 1999). Среднегодовая температура за период 2001–2018 годы составляет 4,9 °C, сумма осадков – 764,9 мм (данные метеостанции «Лесной заповедник», находятся в архиве ЦЛГПБЗ). Луга в заповеднике занимают менее 1 % территории (Кураева и др., 1999). Материковые луга представлены сообществами класса Molinio-Arrhenatheretea R. Tx. 1937 (настоящие луга), но в заповедном ядре они выведены из хозяйственного использования, а в буферной зоне используются как пастбища и сенокосы. Влажные местообитания в понижениях и вдоль русел водотоков заняты сообществами с доминированием таволги вязолистной, относящимися к тому же классу Molinio-Arrhenatheretea. На богатых азотом участках неиспользуемых угодий развиты субрудеральные сообщества класса Epilobietea angustifolii (Cherednichenko, Borodulina, 2018). Пойменные луга относятся преимущественно к классу Phragmito–Magnocaricetea (Чередниченко, 2017).
Эксперименты с разложением стандартного материала. В качестве стандартного материала для оценки скорости разложения ветоши были использованы зеленый чай (EAN: 87 22700 05552 5) и ройбос (EAN: 87 22700 18843 8) марки Lipton в пакетиках-пирамидках из нейлоновой сетки, размер ячейки которой позволяет почвенным микроорганизмам и мезофауне проникать внутрь, а сам пакетик не разлагается во время инкубации. Перед началом работы была определена масса каждого пакетика, включая этикетку, и все образцы
Таблица 1
Характеристика изученных травяных сообществ Звенигородской биостанции (ЗБС) и Центрально-Лесного государственного природного биосферного заповедника (ЦЛГПБЗ) с оценками по экологическим шкалам Элленберга
| Сообщество | Положение в рельефе | Доминирующие виды
(в порядке убывания ПП) |
Союз | Тип использования | F | N | R | |
|
ЗБС |
||||||||
| ПЛ 1 | Низкая пойма | Urtica dioica, Bromopsis inermis, Phalaroides arundinacea | Senecionion fluviatilis | Рекреация | 6,22 | 6,79 | 7,38 | |
| ПЛ 2 | Прирусловой вал | Bromopsis inermis, Artemisia vulgaris, Urtica dioica | Senecionion fluviatilis | Рекреация | 5,57 | 6,69 | 7,27 | |
| ПЛ 3 | Склон высокой поймы | Arrhenatherum elatius, Seseli libanotis | Arrhenatherion elatioris | Рекреация | 5,05 | 4,98 | 7,00 | |
| ПЛ 4 | Склон высокой поймы | Briza media, Carex hirta, Dactylis glomerata, Festuca pratensis, Geum rivale, Seseli libanotis | Arrhenatherion elatioris | Рекреация | 5,09 | 4,73 | 7,52 | |
| ПЛ 5 | Высокая пойма | Arrhenatherum elatius, Geum rivale, Seseli libanotis | Cynosurion cristati | Рекреация, слабый выпас | 5,07 | 5,18 | 7,07 | |
| ПЛ 6 | Высокая пойма | Festuca rubra, Geum rivale | Cynosurion cristati | Рекреация, слабый выпас | 5,33 | 5,02 | 6,76 | |
| ТС 1 | Понижение на высокой пойме | Filipendula ulmaria, Urtica dioica | Filipendulion ulmariae | Не используется | 6,12 | 6,00 | 7,44 | |
| ЦЛГПБЗ | ||||||||
| СЛ 1 | Водораздел | Alchemilla vulgaris, Agrostis tenuis, Anthoxanthum odoratum, Centaurea phrygia, Dactylis glomerata, Hypericum maculatum, Ranunculus acris | Cynosurion cristati | Заповедный режим | 5,24 | 4,01 | 5,02 | |
| СЛ 2 | Водораздел | Centaurea phrygia, Festuca pratensis, Succisa pratensis, Trifolium medium | Cynosurion cristati | Нерегулярный выпас и сенокошение | 5,83 | 4,33 | 5,29 | |
| СРС | Водораздел | Anthriscus sylvestris, Trifolium medium, Chamaenerion angustifolium, Dactylis glomerata, Urtica dioica | Epilobion angustifolii | Заповедный режим | 5,25 | 5,16 | 6,00 | |
| ТС 2 | Понижение на водоразделе | Filipendula ulmaria, Lysimachia vulgaris, Equisetum fluviatile, Ranunculus repens | Filipendulion ulmariae | Не используется | 7,55 | 5,68 | 6,32 | |
Примечание к таблице. Названия союзов приведены в соответствии с Продромусом растительности Европы (Mucina et al., 2016). Латинские названия видов приведены по сводке С. К. Черепанова (1995). Оценки по экологическим шкалам, факторы: R – кислотность; F – увлажнение; N – богатство почвы.
были пронумерованы. Образцы были закопаны в почву на глубину 8 см в пятикратной повторности и располагались на трансекте парами зеленый чай – ройбос, расстояние между парами составило 1 м, а между образцами в паре – 10 см. На ЗБС образцы были заложены 23 июня и извлечены 1 октября 2017 года, инкубационный период составил 108 дней. В ЦЛГПБЗ инкубационный период составил 87 дней (с 11 июля по 5 октября 2019 года). После удаления загрязнений с поверхности образцов, их содержимое извлекали и помещали в бумажные пакеты, предварительно удалив вросшие корни и проникших внутрь беспозвоночных. Образцы высушивали при 80 °С в течение 8 часов и взвешивали с точностью до 0,1 мг, а затем прокаливали при температуре 500 °С в течение 6 часов и взвешивали оставшуюся золу, массу которой затем вычитали из массы образцов, чтобы исключить загрязнение частицами почвы.
Для каждого образца был рассчитан процент потери массы. Так как время инкубации на ЗБС и в ЦЛГПБЗ было разным, то потери массы на ЗБС с более длительным периодом инкубации в 108 дней были скорректированы для срока 87 дней, чтобы можно было сравнивать скорости разложения между сообществами в двух регионах. Для этого по экспоненциальной модели были определены константы скорости разложения каждого образца, и с их использованием потери массы были пересчитаны для периода 87 дней. Значения фактора стабилизации STBI и константы скорости разложения лабильной фракции kTBI были определены по методике, описанной в работе Й. А. Кеускампа с соавторами (Keuskamp et al., 2013). Фактор стабилизации STBI был рассчитан по формуле S=1-ag/Hg, где ag – отношение между потерей массы и исходной массой образца зеленого чая, а Hg – стандартная величина гидролизуемой (лабильной) фракции зеленого чая данной марки, равная 0,842 г/г. Далее мы оценили величину лабильной фракции ройбоса (ar) с учетом фактора стабилизации в данном местообитании по формуле ar=Hr(1-S), где Hr – стандартная величина гидролизуемой фракции ройбоса, равная 0,552 г/г (Keuskamp et al., 2013). Таким образом, масса гидролизуемой фракции, не разложившейся за период экспозиции, для каждого образца ройбоса ar(t) была рассчитана как ar(t)=Wr(t)-(1-ar), где Wr(t) – масса образца ройбоса после окончания опыта. Полученные значения ar и ar(t) были использованы для вычисления kTBI с помощью экспоненциальных моделей разложения для каждого образца ройбоса.
Характеристика факторов среды. В каждом сообществе непосредственно на площадке были выполнены геоботанические описания (25 м2 для сообществ ЗБС и 100 м2 для ЦЛГПБЗ) и по экологическим шкалам Х. Элленберга (Ellenberg et al., 1991) в программе EcoScale v.5 (Грохлина, Ханина, 2015) рассчитаны индикаторные значения для влажности, кислотности и богатства почвы азотом.
Статистическая обработка. Обработка материала проведена в статистической среде R (R Core Team, 2015). Для выявления различий между сообществами по потере массы зеленым чаем и ройбосом, STBI и kTBI был проведен непараметрический дисперсионный анализ Краскела-Уоллиса (Kruskal-Wallis ANOVA) с последующим апостериорным тестом Тьюки-Крамера-Неменьи в пакете PMCMR (Pohler, 2014). Распределение средних значений STBI и kTBI по сообществам не отличалось от нормального, и для выявления связи между ними использовали коэффициент корреляции Пирсона.
Для того чтобы выявить, как параметры разложения связаны с индикаторными значениями влажности, кислотности и богатства почв, были использованы линейные регрессионные модели со смешанным эффектом (Mixed Effect Models, пакет nlme (Pinheiro et al., 2015)), где зависимыми переменными были потери массы зеленым чаем и ройбосом (для ЗБС в пересчете на 87 дней), STBI и kTBI, независимыми фиксированными эффектами были индикаторные значения (количественные) и район исследования (качественный), а случайным эффектом было сообщество, чтобы учесть пространственную неоднородность. Отбор наиболее значимых факторов проводили автоматически с помощью функции stepAIC. Значимость факторов оценивали с помощью дисперсионного анализа II типа (функция «Anova» в пакете car (Fox, Weisberg, 2011)). Качество полученных моделей оценивали по графикам квантиль-квантильного распределения, при необходимости зависимая переменная была логарифмирована по основанию натурального логарифма. Коэффициент детерминации R2 был рассчитан по алгоритму (Nakagawa, Schielzeth, 2013) в пакете MuMIn (Bartoń, 2017]. В модели, где зависимой переменной была kTBI, не вошло сообщество прируслового вала ЗБС, где за период инкубации в четырех из пяти образцов лабильной фракции ройбоса не осталось: часть ее перешла в стабильное состояние (ar оказалось значительно меньше Hr), а другая часть разложилось, и рассчитать kTBI по методике Й. А. Кеускампа с соавторами (Keuskamp et al., 2013) было невозможно.
РЕЗУЛЬТАТЫ
Градиент факторов среды по экологическим шкалам включал сообщества в диапазоне от 5,0 до 7,5 баллов по влажности, от 4,0 до 6,8 по богатству почвы и от 5,0 до 7,5 по кислотности (табл. 1). Индикаторные значения кислотности и богатства были между собой значимо скоррелированы (Spearman R=0,668, p=0,025), при этом средние баллы богатства не отличались между регионами, а в пойменных сообществах ЗБС почвы были менее кислые (p=0,006).
Таблица 2
Параметры разложения стандартного материала – зеленого чая и ройбоса (среднее ± ошибка среднего)
| Сообщество | Потери массы зеленым чаем, % | Потери массы ройбосом, % | Фактор стабилизации STBI, г/г | Константа разложения kTBI, г/г/день |
| ПЛ1 | 52,3±1,9a | 32,7±2,0ac | 0,379±0,022a | 0,0259±0,0029a |
| ПЛ2 | 53,2±1,2a | 36,6±0,8a | 0,368±0,015a | 0,0400 |
| ПЛ3 | 54,3±1,2a | 35,6±1,4ac | 0,355±0,015a | 0,0308±0,0026a |
| ПЛ4 | 57,2±0,8a | 34,6±1,7ac | 0,320±0,010a | 0,0214±0,0010ab |
| ПЛ5 | 65,9±1,9ab | 30,7±1,3ab | 0,217±0,022ab | 0,0158±0,0018ab |
| ПЛ6 | 63,3±3,3ab | 28,0±1,8ab | 0,249±0,039ab | 0,0151±0,0027ab |
| ТС1 | 62,2±1,0ab | 36,3±2,0ab | 0,261±0,012ab | 0,0239±0,0035a |
| СЛ1 | 65,3±0,9ab | 26,8±1,2bc | 0,225±0,010ab | 0,0115±0,0010ab |
| СЛ2 | 65,2±0,9ab | 30,2±1,6ab | 0,226±0,010ab | 0,0149±0,0015ab |
| CРС | 58,4±2,6ab | 29,3±1,2ab | 0,306±0,031ab | 0,0156±0,0018ab |
| ТС2 | 74,0±1,5b | 25,8±0,8b | 0,122±0,018b | 0,0089±0,0004b |
Примечание к таблице. Данные для ЗБС пересчитаны на период 87 дней. Повторность составила n=5 для всех случаев потери массы зеленым чаем и для фактора стабилизации, кроме используемого суходольного луга (СЛ2), где n=4; n=5 для потери массы ройбосом, кроме субрудерального сообщества (СРС), где n=4; повторность для константы разложения: n=5 в СЛ1, СЛ2, ПЛ5, ПЛ6; n=4 в CРС, ТС1 и ТС2; n=3 в ПЛ1, ПЛ3, ПЛ4; n=1 в ПЛ2. Разными буквами отмечены значимые отличия между сообществами (p<0,05).
Максимальные потери массы зеленым чаем за 87 дней отмечены для таволгового сообщества в ЦЛГПБЗ (ТС2) и составили в среднем 74,0 %, а минимальные – для сообществ низкой части поймы и прируслового вала (52,3 % и 53,2 %; табл. 3). Максимальные потери массы ройбосом наблюдались на прирусловом валу (36,6 %) и в пойменном таволговом сообществе ЗБС (36,3 %). В сообществах низкой части поймы, прируслового вала и склона центральной части поймы для нескольких образцов эти потери превысили расчетную массу лабильной фракции (Keuskamp et al., 2013). Медленнее всего образцы ройбоса разлагались в почве таволгового сообщества ЦЛГПБЗ (ТС2; 25,8 %). Крайние значения потерь массы обоих видов значимо отличались друг от друга.
Значения STBI варьировались от 0,122 (таволговое сообщество ЦЛГПБЗ, ТС2) до 0,379 (прирусловой вал), а kTBI – от 0,0089 в таволговом сообществе ЦЛГПБЗ до 0,0308 на склоне центральной части поймы с райграсом и более 0,04 на прирусловом валу, где у четырех из пяти образцов полностью разложилась лабильная фракция ройбоса. Средние значения STBI и kTBI были положительно скоррелированы (r=0,824, p=0,002; рис. 1).
Индикаторные оценки кислотности почвы оказались лучшими предикторами потерь массы ройбосом: на кислых почвах разложение шло медленнее (рис. 2а). Эта связь значима
Таблица 3
Связь параметров разложения стандартного материала (зеленый чай и ройбос) с регионом и характеристиками местообитания
| Материал | Фактор | n | Эффект | χ2 | p | R2m | R2c |
| Потери массы ройбосом | R | 11 | + | 10,92 | <0,001 | 0,315 | 0,523 |
| Потери массы ройбосом (только для ЗБС) | R | 7 | + | 5,80 | 0,016 | 0,221 | 0,350 |
| Фактор стабилизации STBI | F
N |
11 | –
+ |
19,04
15,76 |
<0,001
<0,001 |
0,535 | 0,716 |
| Константа разложения kTBI | Регион | 10 | ЗБС>ЦЛГПБЗ | 6,13 | 0,002 | 0,384 | 0,677 |
| Константа разложения kTBI | R | 10 | + | 6,13 | 0,013 | 0,310 | 0,710 |
Примечание к таблице. Оценки по экологическим шкалам: R – кислотность; F – увлажнение; N – богатство почвы. n – число сообществ, включенных в анализ. Эффекты: «+» – положительная связь; «–» – отрицательная связь. R2m – коэффициент детерминации только для фиксированных факторов; R2c – коэффициент детерминации для фиксированных и случайных эффектов.
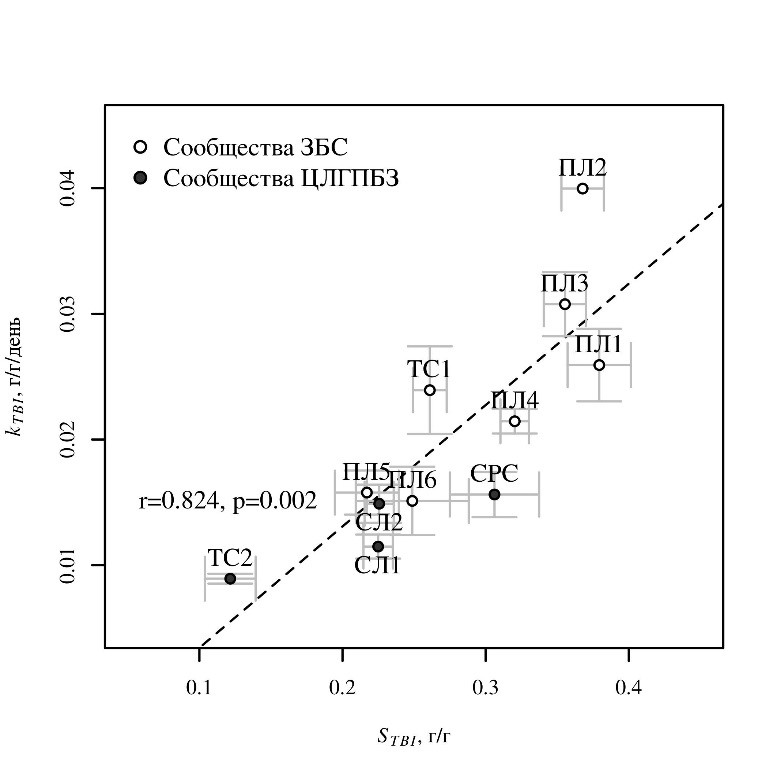
Рис. 1. Связь фактора стабилизации STBI и константы разложения kTBI в травяных сообществах ЦЛГПБЗ и ЗБС
Показаны средние значения и ошибка среднего. Сообщества: Союз Senecionion fluviatilis: ПЛ1 – луг в низкой пойме; ПЛ2 – луг на прирусловом валу. Союз Arrhenatherion elatioris: ПЛ3, ПЛ4 – луга на склоне высокой поймы. Союз Cynosurion cristati: ПЛ5, ПЛ6 – луга в высокой пойме; СЛ1, СЛ2 – луга на водоразделе. СРС – субрудеральное сообщество союза Epilobion angustifolii. ТС1, ТС2 – таволговые сообщества союза Filipendulion ulmariae. Названия союзов приведены в соответствии с Продромусом растительности Европы (Mucina et al., 2016).
как для всех сообществ, так и только для сообществ ЗБС. Константа разложения также была значимо связана с кислотностью почвы (рис. 2б), но при этом лучшей моделью оказалась та, где из предикторов в результате автоматической пошаговой регрессии остался только регион. Таким образом, в пойменных сообществах ЗБС значения kTBI были выше, чем в ЦЛГПБЗ.
Лучшая модель для STBI включала индикаторные значения влажности и богатства почвы (табл. 3): его значения были ниже на влажных и богатых почвах.
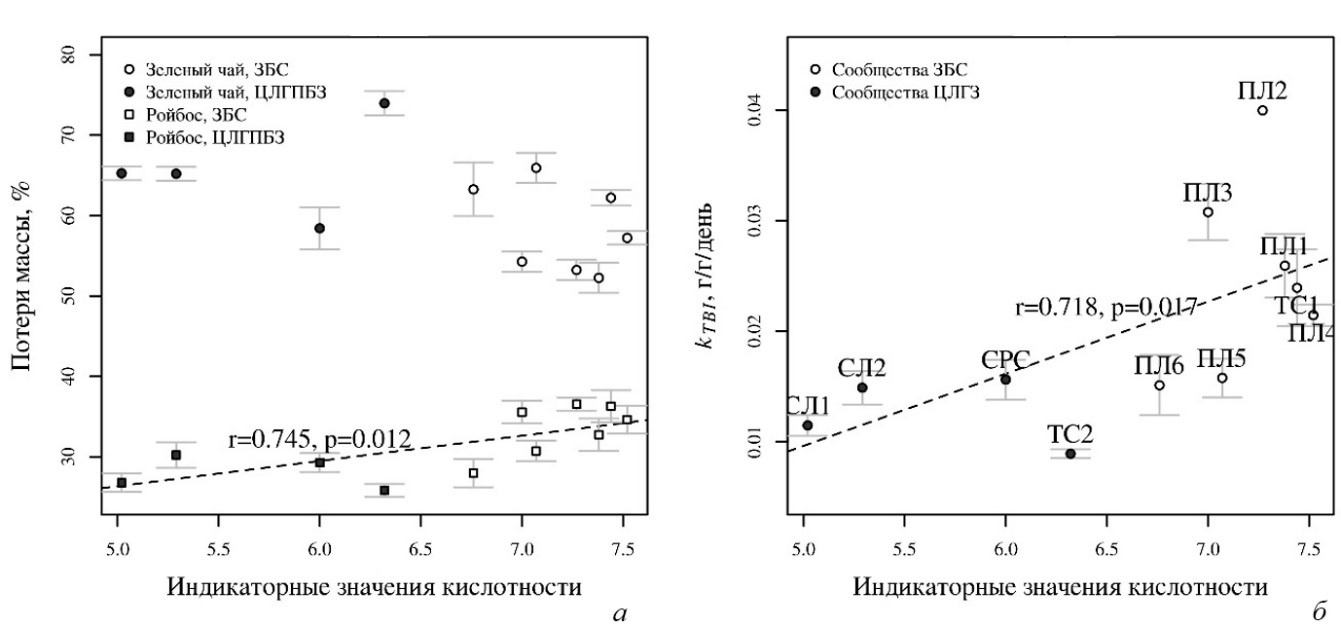
Рис. 2. Связь потери массы зеленым чаем и ройбосом (а) и константы скорости разложения kTBI c индикаторными оценками кислотности почвы (б)
r – коэффициент корреляции Спирмена для средних значений по сообществам. Показаны средние значения и ошибка среднего. Сообщества: Союз Senecionion fluviatilis: ПЛ1 – луг в низкой пойме; ПЛ2 – луг на прирусловом валу. Союз Arrhenatherion elatioris: ПЛ3, ПЛ4 – луга на склоне высокой поймы. Союз Cynosurion cristati: ПЛ5, ПЛ6 – луга в высокой пойме; СЛ1, СЛ2 – луга на водоразделе. СРС – субрудеральное сообщество союза Epilobion angustifolii. ТС1, ТС2 – таволговые сообщества союза Filipendulion ulmariae. Названия союзов приведены в соответствии с Продромусом растительности Европы (Mucina et al., 2016).
ОБСУЖДЕНИЕ
Почвы под травяными сообществами Средней России разнообразны по параметрам разложения, что было показано даже на небольшом числе местообитаний. Так как ЦЛГПБЗ и ЗБС располагаются на значительном расстоянии друг от друга, то ожидалось, что в первую очередь на параметры разложения стандартного материала будет влиять разница в климате, что было показано в ряде работ (Petraglia et al., 2019). Кроме того, наблюдения проводились в разные годы, и разница между сообществами в разных регионах может объясняться погодными условиями. На ЗБС разложение шло при более теплой и влажной погоде: там за период инкубации в 2017 году средняя температура воздуха составила 15,5 ºС, а сумма осадков – 508,4 мм (http://rp5.ru, метеостанция Ново-Иерусалим, ближайшая к ЗБС); а за период инкубации в ЦЛГПБЗ в 2019 году средняя температура была 12,0 ºС, а сумма осадков – 396,1 мм (Шуйская, 2019). Действительно, константа разложения на ЗБС оказалась значимо выше, то есть разложение лабильной фракции ройбоса шло быстрее. Также почвы травяных сообществ ЦЛГПБЗ более кислые, чем в пойме реки Москвы на ЗБС, и связь потери массы ройбосом и kTBI с индикаторными значениями кислотности может быть отражением разницы между почвенными условиями в регионах. Так, значимая связь потери массы ройбосом при включении в анализ только пойменных сообществ ЗБС позволяет предположить, что кислотность почвы в ряду травяных сообществ действительно является важным фактором, определяющим скорость разложения органического вещества, а наблюдаемая разница в температурном режиме не является определяющей.
Кроме того, следует отметить высокое сходство в параметрах разложения стандартного материала между сообществами союза Cynosurion cristati: суходольными лугами ЦЛГПБЗ и не затапливаемыми лугами высокой поймы (фактически по режиму также суходольными лугами) ЗБС (табл. 1, рис. 1). Их значения STBI и kTBI близки к отмеченным на приморских лугах в Нидерландах, где на лугах без распашки в течение 5–10 лет и в течение более 20 лет STBI составлял в среднем 0,224 и 0,208 соответственно, а kTBI – 0,0158 и 0,0165, при этом возраст луга и STBI были отрицательно скоррелированы (Iepema et al., 2015).
В серии изученных травяных сообществ Средней России STBI и kTBI были положительно скоррелированы (рис. 1). В альпийских сообществах Тебердинского заповедника связи между этими двумя величинами выявлено не было (Elumeeva et al., 2018). Это связано с тем, что высокие значения STBI в пойменных сообществах ЗБС могут быть обусловлены высоким содержанием в почвенных растворах кальция, вымываемого рекой из карбонатных пород: 85 мг/л на лугах высокой поймы и 148 мг/л в низкой пойме при pH>8 (Чередниченко, Первова, 2016). В таких условиях стабилизация органического вещества в составе органо-минеральных соединений идет преимущественно за счет кальция, в то время как в кислых почвах – за счет алюминия (Rowley et al., 2018). Возможно, это одна из причин отсутствия значимых различий между значениями STBI на основных почвах поймы реки Москвы и на кислых почвах ЦЛГПБЗ с pH 4,3–5,3 (неопубликованные данные). Также низкие значения STBI характерны для влажных богатых почв под таволговыми сообществами. Повышенная влажность обуславливает наличие анаэробных участков, недоступных для микроорганизмов, а добавление азота усиливает образование гумуса (Prescott, 2010), поэтому в таволговых сообществах можно было бы ожидать высокого уровня стабилизации. С другой стороны, снижение фактора стабилизации при увеличении обводненности и богатства было также отмечено и для прибрежных засоленных местообитаний (Mueller et al., 2018). По-видимому, такие условия важны для сохранения органического вещества в составе трудно разлагаемых фракций, а в случае лабильной фракции способствуют более быстрому ее разложению и/или вымыванию.
В целом, роль факторов среды в определении параметров разложения варьируется по местообитаниям. Так, в пойменных лесных сообществах Квебека (Канада) ни STBI, ни kTBI не зависели от свойств почвы (Saint-Laurent, Arsenault-Boucher, 2020). Отрицательная корреляция STBI с pH почвы отмечена для болот в Канаде (McDonald et al., 2018) и для альпийских сообществ Тебердинского заповедника (Elumeeva et al., 2018), в то время как значимых связей с pH для kTBI не наблюдалось. В ряду травяных сообществ Средней России, напротив, с кислотностью почвы были связаны kTBI и потери массы ройбосом (рис. 2а и 2б). Это согласуется с представлениями о том, что на почвах с pH близким к нейтральному разложение идет быстрее из-за более активной деятельности микроорганизмов (Rowley et al., 2018). Известно, что скорость разложения легко разлагаемого растительного материала сильнее зависит от климатических факторов, а трудно разлагаемого – от свойств почвы и состава растительности (Fanin et al., 2020). В нашем случае градиент по климату относительно небольшой, поэтому разница в скорости разложения ройбоса выражена сильнее и определяется свойствами почвы и произрастающим на ней травяным сообществом.
ЗАКЛЮЧЕНИЕ
Изученные травяные сообщества Средней России оказались разнообразными по параметрам разложения стандартного материала, рассчитанным на основе индекса TBI. Скорость разложения была ниже в сообществах ЦЛГПБЗ и была положительно скоррелирована с индикаторными значениями кислотности почвы по шкале Элленберга. Значения фактора стабилизации не зависели от региона и снижались при увеличении влажности и богатства почвы. Суходольные луга ЦЛГПБЗ и луга высокой поймы реки Москвы, относящиеся к союзу Cynosurion cristati, оказались наиболее сходными по параметрам разложения.
Благодарности. Авторы выражают благодарность А. С. Желтухину, заместителю директора по науке Центрально-Лесного государственного природного биосферного заповедника за возможность проведения полевых работ.
Полевые работы в Центрально-Лесном заповеднике, обработка материала и написание статьи выполнены при поддержке гранта РФФИ № 19-04-00799. Полевые работы на Звенигородской биостанции в 2017 году проведены в рамках государственного задания МГУ № 01201157316 по теме «Механизмы структурно-функциональной организации растительного покрова и рациональное природопользование».
REFERENCES
Александровский А. Л., Панин А. В. Закономерности формирования поймы реки Москвы в контексте ее освоения человеком (на примере ключевого участка в районе Звенигородской биостанции МГУ) // Археология Подмосковья. – 2013. – Т. 9. – С. 249–256.
Грохлина Т. И., Ханина Л. Г. О компьютерной обработке геоботанических описаний по экологическим шкалам // Математическое моделирование в экологии: Материалы Четвертой Национальной науч. конф. с международным участием (Пущино, 18-22 мая 2015 г.). – Пущино: ИФХиБПП РАН, 2015. – С. 63–64.
Копцик Г. Н., Рыжова И. М. Почвы Звенигородской биостанции. – Руководство по летней учебной практике студентов-биологов на Звенигородской биостанции им. С. Н. Скадовского. – М.: Изд-во МГУ, 2011. – С. 14–58.
Кураева Е. Н., Минаева Т. Ю., Шапошников Е. С. Типологическая структура и флористическое разнообразие лесных сообществ // Сукцессионные процессы в заповедниках России и проблемы сохранения биологического разнообразия / [Ред. Смирнова О. В., Шапошников Е. С.]. – Санкт-Петербург: РБО, 1999. – С. 314–323.
Минаева Т. Ю., Шапошников Е. С. Характеристика региона и природные условия территории // Сукцессионные процессы в заповедниках России и проблемы сохранения биологического разнообразия / [Ред. О. В. Смирнова, Е. С. Шапошников]. – Санкт-Петербург: РБО, 1999. – С. 296–299.
Погода и климат [Электронный ресурс]. – http://www.pogodaiklimat.ru/history/27511.htm (дата обращения 16.06.2020).
Чередниченко О. В. Флористическая классификация гигрофитной растительности в местах поселений бобров в пойме реки Межа (Центрально-Лесной заповедник, Тверская область) // Бюллетень Брянского отделения Русского ботанического общества. – 2017. – Т. 9, № 1. – С. 37–49.
Чередниченко О. В., Первова Н. Е. Экологические особенности луговых биогеоценозов на территории Звенигородской биостанции МГУ // АгроЭкоИнфо, 2016. – № 4. – http://agroecoinfo.narod.ru/journal/STATYI/2016/4/st_452.doc
Чередниченко О. В., Ямалов С. М. Растительность пойменных лугов на территории Звенигородской биостанции МГУ // Труды Звенигородской биологической станции им. С. Н. Скадовского. Т. 5 / [Ред. Уланова Н. Г., Гаврилов В. М.]. – М.: Изд-во МГУ, 2011. – С. 166–173.
Черепанов С. К. Сосудистые растения России и сопредельных государств. – Cанкт-Петербург: Мир и семья, 1995. – 992 с.
Шуйская Е. А. Метеорологические характеристики сезонов // Динамика явлений и процессов в природном комплексе заповедника. Летопись природы Центрально-Лесного заповедника, 2019. – Т. 58 (2018). – С. 22–41.
Bartoń K. MuMIn: Multi-Model Inference. R package version 1.40.0, 2017. – https://CRAN.R-project.org/package=MuMIn
Cherednichenko O., Borodulina V. Biodiversity of herbaceous vegetation in abandoned and managed sites under protection regime: A case study in the Central Forest Reserve, NW Russia // Hacquetia. – 2018. – Vol. 17. – P. 35–60.
Ellenberg H., Weber H. E., Dull R. et al. Zeigerwerte Von Pflanzen In Mitteleuropa [Indicator Values of Plants in Central Europe] // Scripta Geobotanica. Vol. 18. – Gottingen: Verlag Erich Goltze Kg, 1991. – 248 s.
Elumeeva T. G., Onipchenko V. G., Akhmetzhanova A. A., Makarov M. I. Stabilization versus decomposition in alpine ecosystems of the Northwestern Caucasus: the results of a tea bag burial experiment // Journal of Mountain Science. – 2018. – Vol. 15, N 8. – P. 1633–1641.
Fanin N., Bezaud S., Sarneel J. M., Cecchini S., Nicolas M., Augusto L. Relative importance of climate, soil and plant functional traits during the early decomposition stage of standardized litter // Ecosystems. – 2020. – Vol. 23, N 5. – P. 1004–1007.
Fox J., Weisberg S. An {R} Companion to Applied Regression, Second Edition. Sage: Thousand Oaks CA, 2011. – URL: https://socserv.socsci.mcmaster.ca/jfox/Books/Companion
Freschet G. T., Aerts R., Cornelissen J. H. C. A plant economics spectrum of litter decomposability // Functional Ecology. – 2012. – Vol. 26, N 1. – P. 56–65.
Fujii S., Mori A. S., Koide D., Makoto K., Matsuoka S., Osono T., Forest I. Disentangling relationships between plant diversity and decomposition processes under forest restoration // Journal of Applied Ecology. – 2017. – Vol. 54, N 1. – P. 80–90.
Iepema G. L., Domhof B., Van Eekeren N. Capacity of the soil to decompose organic matter in old and young grasslands // Grassland science in Europe. Vol. 20. Grasslands and forages in high output dairy farming systems / [Eds. van den Pol-van Dasselaar A. et al.], 2015. – P. 434–436.
Keuskamp J. A., Dingemans B. J. J., Lehtinen T., Sarneel J. M., Hefting M. M. Tea Bag Index: a novel approach to collect uniform decomposition data across ecosystems // Methods in Ecology and Evolution. – 2013. – Vol. 4, N 11. – P. 1070–1075.
MacDonald E., Brummel M. E., Bieniada A., Elliott J., Engering A., Gauthier T.-L., Saraswati S., Touchette S., Turmel-Courchesne L., Strack M. Using the Tea Bag Index to characterize decomposition rates in restored petlands // Boreal Environment Research. – 2018. – Vol. 23. – P. 221–235.
Mucina L., Bültmann H., Dierßen K., Theurillat J.-P., Raus T., Čarni A., Šumberová K., Willner W., Dengler J., Gavilán García R., Chytrý M., Hájek M., Di Pietro R., Iakushenko D., Pallas J., Daniëls F. J. A., Bergmeier E., Santos Guerra A., Ermakov N., Valachovič M., Schaminée J. H. H., Lysenko T., Didukh Y. P., Pignatti S., Rodwell J. S., Capelo J., Weber H. E., Solomeshch A., Dimopoulos P., Aguiar C., Hennekens S. M., Tichý L. Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities // Applied Vegetation Science. – 2016. – Vol. 19, Suppl. 1. – P. 3–264.
Mueller P., Schile-Beers L. M., Mozdzer T. J., Chmura G. L., Dinter T., Kuzyakov Y., de Groot A. V., Esselink P., Smit C., D’Alpaos A., Ibáñez C., Lazarus M., Neumeier U., Johnson B. J., Baldwin A. H., Yarwood S. A., Montemayor D. I., Yang Z., Wu J., Jensen K., Nolte S. Global-change effects on early-stage decomposition processes in tidal wetlands – implications from a global survey using standardized litter // Biogeosciences. – 2018. – Vol. 15, N 10. – P. 3189–3202.
Nakagawa S., Schielzeth H. A general and simple method for obtaining R2 from generalized linear mixed-effects models // Methods in Ecology and Evolution. – 2013. – Vol. 4, N 2. – P. 133–142.
Petraglia A., Cacciatori C., Chelli S. Fenu G., Calderisi G., Gargano D., Abeli T., Orsenigo S., Carboniani M. Litter decomposition: effects of temperature driven by soil moisture and vegetation type // Plant and Soil. – 2019. – Vol. 435, N 1–2. – P. 187–200.
Pinheiro J., Bates D., DebRoy S. Sarkar D., R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-121. 2015. – <URL: http://CRAN.R-project.org/package=nlme>
Pohler T. The Pairwise Multiple Comparison of Mean Ranks Package (PMCMR). – R package, 2014.
Prescott C. E. Litter decomposition: what controls it and how we can alter it to sequester more carbon to forest soils? // Biogeochemistry. – 2010. – Vol. 101, N 1–3. – P. 133–149.
R Core Team R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2015. – URL https://www.R-project.org/
Rowley M. C., Grand S., Verrecchia E. Calcium-mediated stabilization of soil organic carbon // Biogeochemistry. – 2018. – Vol. 137. – P. 27–49.
Saint-Laurent D., Arsenault-Boucher L. Soil properties and rate of organic matter decomposition in riparian woodlands using the TBI protocol // Geoderma. – 2020. – Vol. 358, N 1–2. – 113976. – P. 1–10.
Stockmann U., Adams M. A., Crawford J. W., Field D. J., Henakaarchchi N., Jenkins M., Minasny B., McBratney A. B., de Remy de Courcelles V., Singh K., Wheeler I., Abbott L., Angers D. A., Baldock J., Bird M., Brookes P. C., Chenu C., Jastrow J. D., Lal R., Lehmann J., O’Donnell A. G., Parton W. J., Whitehead D., Zimmermann M. The knowns, known unknowns and unknowns of sequestration of soil organic carbon // Agriculture, Ecosystems and Environment. – 2013. – Vol. 164, N 1. – P. 80–99.
Whigham D. F., Walker C. M., Maurer J., King R. S., Hauser W., Baird S., Keuskamp J. A., Neale P. J. Watershed influences on the structure and function of riparian wetlands associated with headwater streams – Kenai Peninsula, Alaska // Science of the Total Environment. – 2017. – Vol. 599-600. – P. 124–134.
