
FEATURES OF INTRODUCED SPECIES OF LILAC (SYRINGA) FRUITING IN THE BASHKIR URALS
JOURNAL: «EKOSISTEMY», Issue 22, 2020
Publication text (PDF):Download
UDK: 630*181.522:582.931.4 (470.57-25)
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Polyakova N.V., South-Ural Botanical Garden-Institute of the Ufa Federal Research Centre of the Russian Academy of Sciences, Ufa, Republic of Bashkortostan, Russia
TYPE: Article
DOI:https://doi.org/10.37279/2414-4738-2020-22-90-96
PAGES: from 90 to 96
STATUS: Published
LANGUAGE: Russian
KEYWORDS: Syringa L., fruiting, introduction, Ufa, Bashkir Cis-Urals
ABSTRACT (ENGLISH):
Fruiting is an important characteristic of plant adaptation to new growing conditions. This is especially actual for ornamental plants, which, after successful introduction, can be used in landscaping of settlements. The genus Syringa L. has about 30 species, many of which can replenish the range of ornamental plants in the region. The collection of lilacs of the South Ural Botanical Garden-Institute of the UFIC RAS (Ufa Federal Research Centre of Russian Academy of Sciences) currently contains 11 species that have reached a generative state. The aim of this research is to identify the features of fruiting species of the genus Syringa during introduction in the conditions of the Bashkir Cis-Urals. The fruiting level of lilac species in the collection of the Botanical Garden was studied by determining the total number of fruits of the mother bushes. It is revealed that the fruiting level of introduced species of lilac is determined by the biological (hereditary) characteristics of the species and seasonal changes of weather conditions. S. velutina is characterized by the highest fruiting level, S. vulgaris and S. wolfii – by the lowest ones. It is assumed that there might be a tendency for dependence of the degree of fruiting on the quality of the formed pollen. Two taxa among all studied ones demonstrate fruiting level instability: S. reticulata ssp. reticulata blooms and bears fruit irregularly, while S. reticulata ssp. amurensis has regular abundant flowering, but there is a lack of fruiting in some years.
ВВЕДЕНИЕ
Одной из основных задач ботанических садов является сбор и сохранение коллекций растений, интродуцированных в новой для них климатической зоне, а также долгосрочный мониторинг всех жизненных циклов этих растений. Другая немаловажная задача ботанических садов – разработка рекомендаций по обогащению ассортимента декоративных растений для озеленения мегаполисов и других населенных пунктов конкретного региона. Сирень является одним из наиболее популярных декоративных кустарников, используемых в озеленении (Окунева, 2019). Однако при озеленении городов обычно используют 1–2 вида сирени. Поэтому в сложившейся ситуации актуальным становится пополнение ассортимента не менее декоративными, неприхотливыми и устойчивыми видами сирени. При отборе перспективных видов в целях озеленения необходимо получить комплексную оценку устойчивости этих видов в регионе. Особенности плодоношения являются важной характеристикой адаптированности интродуцентов к новым условиям произрастания и эти показатели используются для оценки перспективности их интродукции (Лапин, Сиднева, 1973; Трулевич, 1991).
В Южно-Уральском ботаническом саду-институте УФИЦ РАН (г. Уфа) на протяжении многих лет проводятся интродукционные исследования родовых комплексов декоративных древесных растений (Мурзабулатова, Полякова, 2015; Мурзабулатова, Полякова, 2016; Мурзабулатова и др., 2018). Коллекция сирени является одной из наиболее крупных в ботаническом саду и на основе ее с начала 2000-х годов проводится комплексное интродукционное изучение представителей рода Syringa (Полякова, 2010, 2016).
Цель данной работы – выявить особенности плодоношения видов сирени при интродукции в условиях Башкирского Предуралья как одного из важнейших показателей интродукционной устойчивости.
МАТЕРИАЛ И МЕТОДЫ
Объектами исследований явились 11 видов рода Syringa коллекции Южно-Уральского ботанического сада-института УФИЦ РАН, достигших генеративного состояния (табл. 1). Все они выращены из семян, полученных в конце 50-х – начале 60-х годов прошлого века из разных ботанических учреждений по программе обмена растительным материалом (Делектус). Все виды в коллекции произрастают в сирингарии, причем виды сосредоточены в начале коллекционного участка.
Климат Башкирского Предуралья, в пределах которого находится ботанический сад, континентальный и характеризуется холодной зимой и теплым летом, большой амплитудой колебаний температуры в течение года, ранними осенними и поздними весенними заморозками.
В Уфе среднемноголетняя температура воздуха составляет +3,4 °C, средняя температура января –13,5 °С, абсолютный минимум достигает –48,5 °С, зимой нередки оттепели, безморозный период длится в среднем 144 дня. Средняя температура июля +19,5 °C, абсолютный максимум зафиксирован на уровне +40 °С. Климат достаточно влажный: среднегодовое количество осадков составляет 500–590 мм, в том числе около 350 мм в теплый период (максимум приходится на июнь – июль). Снежный покров устанавливается в ноябре и держится в среднем 155 суток (Путенихин и др., 2017).
Таблица 1
Характеристика интродуцированных видов сирени
| № | Таксон | Географическое происхождение | Место получения | Год получения |
| 1 | Syringa emodi Wall. | Сев.-Зап. Гималаи | г. Ташкент | 1959 |
| 2 | Syringa × henryi Schneid. | гибрид (S. josikaea × S. villosa) | г. Архангельск | 1958 |
| 3 | Syringa josikaea Jacg. | Карпаты, Трансильвания | Местная репродукция | 1960 |
| 4 | Syringa komarowii Schneid. | Сев.-Зап. Китай | г. Москва | 1967 |
| 5 | Syringa pubescens Turcz. | Сев. Китай, Монголия | Германия | 1960 |
| 6 | Syringa reticulata (Blume) H.Hara ssp. amurensis (Rupr.) P.S. Green & M.C. Chang | Приамурье, Приморский край, Сев.- Вост. Китай | г. Минск | 1961 |
| 7 | Syringa reticulata (Blume) H. Hara ssp. reticulata | Япония (о-ва Хондо, Хоккайдо, Хонсю) | Польша | 1959 |
| 8 | Syringa sweginzowii Koehne et Lingelsh. | Китай, Сев. Корея | г. Ленинград | 1960 |
| 9 | Syringa wolfii Schneid | Дальний Восток, Сев.-Вост. Китай, Корея | г. Ленинград | 1960 |
| 10 | Syringa velutina Kom. | Сев. Китай, Корея | Неизвестно | Неизвестно |
| 11 | Syringa vulgaris L. | Зап. и Юж. Румыния, Югославия, Болгария | г. Киев | 1941 |
Учет степени плодоношения маточных кустов сирени проводился по специальной методике (Методические указания…, 1980). Для этого на каждом экземпляре выделялись модельные ветви (по 1 с каждой из сторон света) длиной около 1 м, измерялась длина каждой ветви по ее оси и суммировалась длина побегов всех порядков ветвления (в метрах). На каждой ветви подсчитывалось количество плодов, затем вычислялось количество плодов на 1 погонный метр данной модельной ветви. После чего определялось среднее количество плодов по всем модельным ветвям данного маточного экземпляра.
Общее приблизительное количество плодов маточника вычислялось по формуле:
Q = 10qRlk,
где:
Q – общее количество (шт.) плодов маточника;
q – количество (шт.) плодов на 1 п. м. модельной ветви;
R – средний радиус округленной проекции кроны;
l – длина протяженности зеленой кроны по стволу;
k – густота кроны в баллах (1–3).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Таблица 2
Степень плодоношения некоторых видов рода Syringa за 1 год в коллекции ботанического сада
| Вид | Средний радиус кроны, м | Длина протяженности зеленой кроны по стволу, м | Густота кроны, баллы | Количество плодов на 1 погонный метр, шт. | Общее количество плодов маточника, шт. | Среднее количество плодов по виду, шт. |
| Syringa emodi 1 | 1,1 | 1,9 | 3 | 57,3 | 3593 | 4737 |
| Syringa emodi 2 | 1,1 | 1,8 | 3 | 108,9 | 5881 | |
| Syringa × henryi 1 | 1,4 | 2,4 | 3 | 164,6 | 16592 | 12834 |
| Syringa × henryi 2 | 1,3 | 2,6 | 3 | 89,5 | 9075 | |
| Syringa josikaea 1 | 1,3 | 3,2 | 3 | 96,6 | 12056 | 12392 |
| Syringa josikaea 2 | 1,4 | 3,2 | 3 | 94,7 | 12728 | |
| Syringa sweginzowii 1 | 1,3 | 2,5 | 3 | 68,9 | 6718 | 7427 |
| Syringa zweginzowii 2 | 1,0 | 2,3 | 3 | 117,9 | 8135 | |
| Syringa velutina 1 | 1,5 | 2,6 | 3 | 32,7 | 3826 | 9468 |
| Syringa velutina 2 | 1,2 | 2,8 | 3 | 149,9 | 15110 | |
| Syringa wolfii 1 | 1,1 | 2,5 | 2 | 63,4 | 3487 | 3351 |
| Syringa wolfii 2 | 1,0 | 2,6 | 2 | 61,8 | 3214 | |
| Syringa vulgaris 1 | 2,1 | 3,1 | 3 | 72,3 | 14120 | 10420 |
| Syringa vulgaris 2 | 1,9 | 3,0 | 3 | 39,3 | 6720 |
Было определено общее количество плодов маточников некоторых видов сиреней (по 2 экземпляра каждого вида) и вычислено среднее количество плодов по видам (табл. 2). Виды S. reticulata ssp. amurensis и S. reticulata ssp. reticulata не задействованы в данном случае, так как цветение и плодоношение у них не ежегодное, а учет плодоношения у S. pubescens и S. komarowii в этот раз не удалось провести по субъективным причинам. Установлено, что максимальная продуктивность за 1 год оказалась у S. × henryi и S. josikaea, высокая – у S. vulgaris (рис. 1а), средняя – у S. velutina и S. sweginzowii, минимальная – у S. emodi и S. wolfii.
Из данных, полученных за несколько лет наблюдений за интродуцированными видами сирени (табл. 3), видно, что высокий показатель плодоношения имеет S. velutina – 156,8 шт./пог. м (рис. 1b). Приближаются к нему также S. sweginzowii (рис. 1c), S. pubescens, S. josikaea и S. emodi (рис. 1d). Характер цветения у этих видов практически ежегодно стабильно высокий (рис. 1). Наименьшей степенью плодоношения характеризуются S. reticulata ssp. reticulata – 19,4 шт./пог. м, S. vulgaris – 38,6 шт./пог. м и S. wolfii – 52,3 шт./пог. м.
В отношении видов с минимальной степенью плодоношения необходимо отметить следующее. S. reticulata ssp. reticulata характеризуется не только нерегулярным, но и необильным плодоношением в те годы, когда оно происходит; кроме того, у этого вида в отдельные годы отсутствует и фаза цветения. При изучении жизнеспособности пыльцы оказалось, что процент прорастания пыльцевых зерен у этой сирени сильно колеблется по годам – от 0 до 60 %. Возможно, причиной этого явления могут быть природно-климатические условия Башкирского Предуралья, которые отличаются от таковых на родине данного вида (о-ва Хондо, Хоккайдо, Хонсю). Другой вид с низким плодоношением, S. wolfii, также родом с Дальнего Востока; в наших условиях он цветет и плодоносит ежегодно, однако обилие этих двух фенофаз стабильно невысокое. Кроме того, возможно, разница в климатических условиях этих регионов (Дальний Восток – муссонный климат и Башкортостан – резко континентальный) служит причиной нерегулярного плодоношения дальневосточных сиреней. Вероятно, имеет место и отсутствие насекомых-опылителей, обитающих в естественных условиях произрастания.
Рис. 1. Цветение некоторых видов сирени, интродуцированных в Башкирское Предуралье
a – Syringa vulgaris; b – S. velutina; c – S. sweginzowii; d – S. emodi.
Таблица 3
Характеристика плодоношения интродуцированных видов сирени за 3 года наблюдений
| Вид | Среднее количество плодов на погонный метр ветви, шт. | Пределы, шт. |
| Syringa emodi | 139,9±23,18 | 57,3–285,0 |
| Syringa × henryi | 108,7±23,48 | 63,0–164,6 |
| Syringa josikaea | 121,7±20,26 | 90,0–217,0 |
| Syringa komarowii | 92,3±22,48 | 34,0–137,0 |
| Syringa pubescens | 133,0±45,00 | 88,0–178,0 |
| Syringa reticulata ssp. amurensis | 129,4±21,82 | 85,4–182,2 |
| Syringa reticulata ssp. reticulata | 19,4±1,99 | 16,2–25,5 |
| Syringa sweginzowii | 134,5±31,09 | 57,0–269,0 |
| Syringa velutina | 156,8±23,78 | 32,7–247,0 |
| Syringa vulgaris | 38,6±10,88 | 4,0–72,3 |
| Syringa wolfii | 52,3±5,32 | 30,0–69,0 |
Продуктивность S. vulgaris также имеет свои особенности. При общем обильном цветении на протяжении многих лет (рис. 1) завязывание плодов у этого вида остается низким, что подтверждается подсчитанным средним количеством плодов на 1 погонный метр модельной ветви. И даже если количество плодов на отдельных модельных ветвях оказывается велико, большинство из них очень мелкие и семена в них либо низкого качества, либо отсутствуют вообще. Низкое качество семян у этого вида подтверждено опытами по определению лабораторной всхожести в предыдущие годы (Полякова, 2011). Подсчитанная же высокая продуктивность S. vulgaris достигнута за счет крупных размеров маточных кустов. Качество семян у сирени прямо пропорционально качеству пыльцы, а у S. vulgaris оно всегда низкое – максимальный процент жизнеспособности пыльцы достигает 22 %. Отсюда и низкий процент завязывания плодов (рис. 2).
Рис. 2. Плодоношение некоторых видов сирени
а – плодоношение Syringa komarowii; b – плодоношение S. vulgaris, значительная часть цветков не сформировала плоды.
При выявлении связи между жизнеспособностью пыльцы и уровнем плодоношения видов коэффициент корреляции составил r=0,48 при p=0,191, что свидетельствует о его недостоверности. Вероятно, это связано с небольшим объемом выборки. Однако, между рассматриваемыми показателями вероятно существование связи средней степени, что наглядно подтверждает приведенный график (рис. 3). Во всяком случае, можно говорить о той же тенденции, которая была отмечена в отношении всхожести семян: более жизнеспособная пыльца способствует не только формированию семян лучшего качества, но и увеличению количества плодов. Что касается S. wolfii, показатели которой отличаются от таковых у других видов на графике, то в этом случае, возможно, имеет место стабильно низкий уровень цветения данного вида в условиях Республики Башкортостан.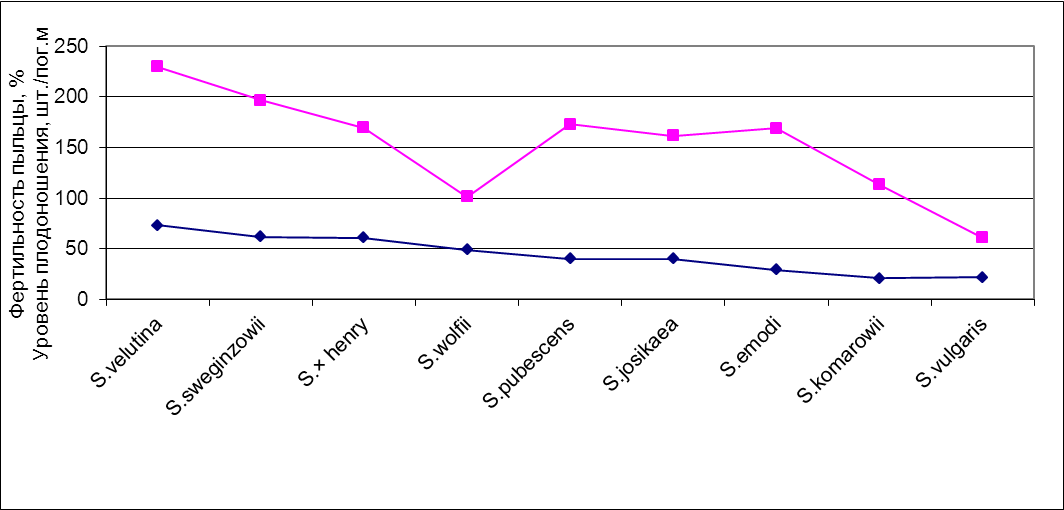
Рис. 3. Связь фертильности пыльцы (нижняя кривая) и уровня плодоношения (верхняя кривая) Виды ранжированы по убыванию фертильности.
Проведенный дисперсионный анализ показал существенное влияние как фактора года (сезонных метеоусловий), так и индивидуальных особенностей видов на уровень плодоношения видов сирени в коллекции (фактор года – F=5344, p<0,001; видовые особенности – F=6011, p=0,0008). Под фактором года имеется в виду совокупность погодных условий – сумма осадков, сумма положительных температур, сумма низких температур в зимний период, высота снежного покрова; под видовым особенностями – сроки наступления фенофаз, особенности морфологии генеративных органов и так далее.
ЗАКЛЮЧЕНИЕ
Таким образом, уровень плодоношения интродуцированных видов сирени в коллекции Южно-Уральского ботанического сада-института Уфимского НЦ РАН определяется биологическими (наследственными) особенностями видов и сезонными изменениями погодных условий. Наибольшим уровнем плодоношения характеризуется Syringa velutina, наименьшим – S. vulgaris и S. wolfii. Возможно, имеет место тенденция зависимости степени плодоношения от качества формируемой пыльцы. Два таксона среди всех изученных обнаруживают нестабильность плодоношения: S. reticulata ssp. reticulata цветет и плодоносит нерегулярно, а у S. reticulata ssp. amurensis при регулярном обильном цветении отмечается отсутствие плодоношения в отдельные годы.
Работа выполнена по Программе фундаментальных исследований Президиума РАН «Биоразнообразие природных систем и биологические ресурсы России» и в рамках государственного задания ЮУБСИ УФИЦ РАН по теме АААА-А18-118011990151-7.
REFERENCES
Лапин П. И., Сиднева С. В. Оценка перспективности интродукции древесных растений по данным визуальных наблюдений // Опыт интродукции древесных растений. – М.: Наука, 1973. – С. 7–67.
Методические указания по семеноведению интродуцентов. – М.: Наука, 1980. – 63 с.
Мурзабулатова Ф. К., Полякова Н. В., Малораспространенные декоративно-лиственные кустарники коллекции Уфимского ботанического сада // «Живые и биокосные системы» [Электронный ресурс]. – 2015. № 13; Режим доступа: http://www.jbks.ru/archive/issue-13/article-5 (просмотрено 16.12. 2019).
Мурзабулатова Ф. К., Полякова Н. В. Интродукция малораспространенных декоративных красивоцветущих кустарников в Башкирском Предуралье // Растительный мир Азиатской России. – 2016. – № 2 (22). – С. 73–83.
Мурзабулатова Ф. К., Полякова Н. В., Никитина Л. С., Путенихин В. П., Шигапов З. Х. Красивоцветущие и декоративно-лиственные кустарники (Фрутицетум, Сирингарий и некоторые другие коллекционные участки Уфимского ботанического сада). – Уфа, «Мир печати», 2018. – 152 с.
Окунева И. Б. Сирень: Уход, обрезка, размножение, сорта. – Москва: Фитон XXI, 2019. – 288 с.
Полякова Н. В. Биологические особенности представителей рода Syringa L. при интродукции в Башкирском Предуралье: дисс… канд. биол. наук: спец.: 03.02.01. Ботаника. – Уфа: БашГУ, 2010. – 152 с.
Полякова Н. В. Биология семян видов сирени в ботаническом саду г. Уфы // Научные ведомости Белгородского государственного университета. Серия: Естественные науки. – 2011. – Т. 14, № 3–1 (98). – С. 56–60.
Полякова Н.В. Интегральная оценка перспективности видов рода Syringa L. в коллекции Уфимского ботанического сада // Известия Уфимского научного центра РАН. – 2016. – № 3. – С. 70–73.
Путенихин В. П., Путенихина К. В., Шигапов З. Х. Кедр сибирский в Башкирском Предуралье и на Южном Урале: биологические и лесоводственные особенности при интродукции. – Уфа: Башкирская Энциклопедия, 2017. – 248 с.
Трулевич Н. В. Эколого-фитоценотические основы интродукции растений. – М.: Наука, 1991. – 214 с.
